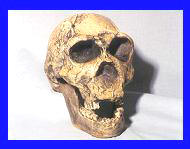
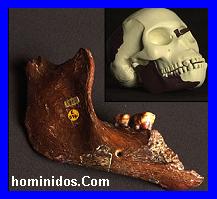
El modelo evolutivo
antropognomònico supone que el proceso filogenético verificado en el
gènero homo, reconoce como motor biológico al crecimiento alomètrico e
hipermòrfico de la capacidad encèfalocraneal, en tanto que su
direccionamiento de cambios se orienta teleológicamente en pos de la
optimización gnomónica del género, mediante la selección natural y
social de los mejor adaptados a los cambios climáticos pleitocènic-holocènicos,
a través de la asimilación, hibridación y/o la sustitución
intersubespecìfica entre todos los ejemplares del gènero homo que
alcanzaron a desarrollarse en cada tiempo y sitio de encuentros
generados por la expansiòn.
Este proceso se
cuantifica a través del cociente de encefalizaciòn y cualitativamente se
verifica en la hipermorfosis craneoencefálica de la evidencia fòsil.
El crecimiento
alomètrico ha sido quien motorizó las modificaciones que se verifican en
las proporciones relativas dentro del propio género y el que originó la
gran divergencia taxonómica que nos separa del resto de los simios
actuales y animales en general, pero por sobre todo, fue también el que
por tener un gradiente de crecimiento temporal y geogràficamente
diferenciado debido a las diferentes condiciones ambientales, habría
sido causa exterior de las distintas subespecies humanas extintas que
evolucionaron de maneras diferentes y divergentes, tan diferentes y
divergentes, como lo fueron las latitudes geográficas y las condiciones
y medios ambientes a que tuvieron que adaptarse lejos de lo que fuera su
cuna africana.
Al parecer habría sido
el homo ergaster el primero en emigrar de Àfrica hace 1,5 millones de
años y en su migrar expansivo se habrían sucedido diversas oleadas del
mismo gènero como asì también de los homo erectus, separadas quizás por
años, muchos años, y siendo cada una de estas oleadas, las portadoras de
su propio estadio de crecimiento hipermòrfico ya iniciado en Àfrica,
como de su tendencia a la verticalizaciòn y gnomizaciòn corporal
creciente, las diferencias de latitudes geográficas y condiciones
medioambientales, fueron sin lugar a dudas, los factores que han marcado
las diferencias de velocidades en el crecimiento alomètrico de cada
grupo homínido y en consecuencia, la causa de las diferenciaciones entre
subespecies del homo.
Así con la
expansión del gènero por Eurasia luego de salir de Àfrica, se
desarrolló la muy difundida figura del candelabro como manera de
explicarnos gráficamente la expansión humana, pero en verdad, no
basta para comprender su complejidad y poligenia, por lo que
debemos poner de relieve e insistir en que la ramificación del
candelabro representando la diferenciación del género, se debe a
las diferencias del gradiente de crecimiento alomètrico impuesto
en cada caso por las condiciones ambientales locales a las que a
cada grupo homínido le tocó adaptarse en latitudes distintas y
que luego, dado el origen del género en las zonas
afro-ecuatoriales y a su tendencia ininterrumpida hacia la
optimización anatómica del cuerpo como gnomon zoológico.
Es
que teleológicamente, la selección entre las subespecies (inter-subespecìfica)
de la evolución poligénica, se orientó dominantemente hacia privilegiar
la adaptación respecto al Sol y fundamentalmente a seleccionar entre los
ejemplares de las subespecies, a los mejor adaptados para valerse
espacial y temporalmente del Sol, del cosmos y sus correlatos con las
migraciones de las presas animales y los alimentos vegetales.
A los más favorecidos
para orientarse por los posicionamientos diarios del sol y las
diferentes estaciones climáticas, ya sea por la conformación gnomónica
del propio cuerpo (la posición bípeda y erecta) como por la experiencia
gnomónica experimental acumulada y transmitida generacionalmente, se le
habrìa facilitado mucho el transitar la geografía e incluso con su
saber, pudieron convencer y arrastrar con ellos a todos aquellos que
desarrollados en latitudes alejadas del eje ecuatorial, no obstante, se
asimilaron mediante un proceso múltiple en el que se habrían conjugado
la inclusión social, la hibridación entre sub-especies diferentes, como
así también la exclusión y sustitución de los componentes.
En la imagen del
candelabro, cada brazo se corresponde con un conjunto de oleadas
migratorias de un taxón determinado que pudo dar lugar a la aparición
poligènica de las subespecies del Homo floresiensis, el Homo
neandertalensis, Homo siberiensis y el Homo que sin salir de su cuna,
siguió evolucionando dentro de Àfrica hasta agotar la totalidad de las
posibilidades del crecimiento alomètrico del encéfalo y su total
consumación hipermòrfica.
Este linaje africano
cuando emigra, lo hace ya totalmente consumado en su anatomía como un
perfecto gnomon zoológico que recto y erguido sobre su bipedismo y a
diferencia de todas las demás especies que lo rodeaban y de las cuales
seguramente se percatò, fuè favorecido por las condiciones del medio
ambiente ecuatorial para entregarse casi como un juego, a la
experimentación con su propio cuerpo y sombra solidaria.
Así surge lo
que decidimos llamar Homo experimentador (OMO I y OMO II) como
el posible primer antropos gnomónico que es anatómicamente un
gnomon zoológico y que como tal experimenta con su sombra pero
que aun no se sabe a/sí mismo un gnomon y que al momento de
salir de Àfrica, quizás ya sabiéndose gnomon, le cupo el
involuntario e inevitable rol de liderar los destinos de la
evolución del gènero, principalmente conduciendo a las especies
que por entonces aùn no habían consumado su evolución
hipermórfica y consiguientemente, de imponer la conducta
gnomònica en relación con los correlatos solares y cosmológicos.
Asì las cosas, el
candelabro se forma por oleadas eruptivas de la diversidad del gènero de
cada época, las que como el magma de un volcán se expandieron sobre la
geografía en torno a su epicentro africano, dando lugar a todas las
variantes biológicas que hoy surgen de la evidencia fòsil.
Dado el carácter
originariamente gnomónico del género homo, las tardías oleadas salientes
de África, posiblemente derivadas del tipo OMO, pudieron liderar
centralmente al conjunto a medida que se encontraban, haciendo que todas
las subespecies se hicieran concurrentes al mismo fin (evolución
teleológica) de alcanzar su óptima relación con el Sol, por lo que sin
proponérselo y de hecho, paulatinamente se ocuparan los mejores sitios
de la franja gnomónica planetaria, donde con el paso del tiempo se verán
florecer las civilizaciones más destacadas de la historia. De esta
manera, el candelabro evolutivo descripto, se pone cabeza abajo por los
entrecruzamientos y por el resultado final de lo que muy vagamente se
suele denominar “el hombre moderno”,
Este hombre moderno es
en un principio un hombre que se sabe productor de sombras propias, y
capaz de servirse de ellas para orientarse, es asì que ese hombre marca
sus tiempos diarios y calendàricos conforme a dichas sombras. Es el
sapiens gnomónico,
Luego con la aparición
del maquinismo, la industria y fundamentalmente el ferrocarril, el
tiempo gnomónico cede su lugar al tiempo mecánico y este al tiempo
electrónico, quebrándose de esta manera el carácter gnomónico del
antropos y convietiendose en un antropos técnico tendiente a la
hibridación completa con la técnica
Conforme a lo descripto,
vemos que el género homo se evidencia como un proceso filogenéticamente
en formación de y manifestaciones diversas no totalmente consumadas en
su crecimiento alomètrico hasta la apariciòn del antropos gnomónico, y
que mediante la selecciòn de los mejor adaptados a las nuevas
condiciones climáticas de finales del pleistoceno y comienzo del
holoceno, se orienta inevitablemente hacia optimizar las relaciones
entre las condiciones anatómicas y subjetivas alcanzadas por el gènero
con el cosmos, tomando al Sol como su principal orientador y referencia.
Los mejores adaptados a
estas condiciones y por lo tanto favorecidos, pudieron ser los
representantes tardíos provenientes de Àfrica.
Este modelo se
funda en que el proceso de crecimiento alomètrico del encéfalo
iniciado en los pre homo hàbilis y su expansión que como
coronación llega a las diferentes subespecies de sapiens/sapiens,
sapiens neandertalensis, sapiens floresiensis, sapiens
siberiensis y sapiens chiniensis, fuè un indetenido proceso
hipermòrfico que una vez iniciado, pudo tener cocientes
incrementales de crecimientos tan diversos como diversos fueron
los medios y condiciones generales impuestas por la expansión y
que tanto ontológica como filogenéticamente, ese proceso de
crecimiento encefálico relativo continuó evolucionando en cada
especie y grado, dentro y fuera de África.
De esta manera, no
habría un hombre moderno como vagamente suele decirse, en todo caso se
verifica que hubo un hombre sabedor de su condición gnomónica que
orientó teleológicamente el proceso evolutivo en lo anatómico y
filogenético (hibridación, sustitución) como en lo social (asimilación,
cruzamientos, asociación) y técnico, dando lugar al desarrollo de las
mas evolucionadas manifestaciones civilizatorias de la antigüedad:
Luego, ese mismo hombre
con experiencia gnomònica, al sustituir el tiempo gnomónico por el
tiempo mecànico, pese a no cambiar anatómicamente pues habría alcanzado
su culminación filogenética, cambia no obstante su imagen y sus modos.
Al igual que otros
modelos de la evolución humana propuestos con anterioridad, el modelo
evolutivo antropognomònico en mérito a las evidencias fósiles y a la
geografìa, concede al continente africano un papel clave en la evolución
de las características modernas y gnomónicas, no obstante, hay que decir
que ese papel clave adjudicado al continente africano, no es lineal ni
absoluto sino que se le da como epicentro de una expansión que en forma
de oleadas eruptivas se difundió a todas las latitudes llevando las
características modernas y gnomónicas adquiridas en África por los
homínidos y humanos que desde allí emigraron tardíamente y que luego,
favorecidos por los cambios climáticos sobre los lugares a que
arribaron, se habrían impuesto y transmitido a través de un proceso
gradual y complejo de asociaciones especìficas e intrespecìficas,,
asimilaciones e hibridaciones por cruzamientos y conflictos diversos.
La ola extendida con
centro en el Este ecuatorial africano, a orillas del Lago Turkana (ex
Rudolf), se extiende por toda Europa y Asia en tanto en Àfrica
continuaron evolucionando los ejemplares kenyanthropus y rudolfensis (en
alusión al Lago Rudolf) que podríamos considerar como muestras concretas
de los primeros pre-antropos gnomònicos o simplemente gnómones
zoológicos en proceso de consumación en función del desarrollo
hipermòrfico y anatòmico
De esta manera los
neandertales y los homos arcaicos que se desarrollaron fuera de Àfrica,
habrìan sido asimilados distintamente por las sucesivas oleadas
eruptivas con centro africano a través de cruzamientos generalizados y
posiblemente también, en torno a la prevaleciente sominaciòn del
carácter gnomónico de la figura bípeda y erecta del africano tardío con
un cerebro capaz de verse a/sì mismo como un objeto fuera de/sì, a la
manera de un cuerpo indisolublemente unido a una sombra que supo
utilizar para orientarse en el espacio y en el tiempo, es decir con
capacidad de ideación simbólica y orientación espaciotemporal que pudo a
su vez generar una tendencia dominante sobre el resto de los humanos en
formación y conducirse geográficamente en pos del Sol hacia un mismo fin
gnomónico y orientador que se verifica en todo lugar donde el sapiens
moderno ha llegado y dejado algún legajo.
En
consecuencia, la evolución y establecimiento de características
modernas y gnomónicas, fue un proceso complejo de mezcla y no
una mecánica y simple sustitución y reemplazo de una especie por
otra. Las evidencias estarían diciendo que se habría dado una
evolución filogenética interespecìfica y teleológicamente
dirigida hacia la consumación del máximo desarrollo encefálico
hipermòrfico correlativamente con la optimización de la
adaptación a las relaciones con el Sol y el cosmos.
Evolución que, con
origen eruptivo en el Este ecuatorial africano, se orientó en pos del
Sol y como resultado final supo arrojar lo que conocemos hoy como
subespecie sapiens/sapiens. Evolución que reabre la discusión en torno
al concepto de especie y que desde lo gnomónico se vislumbra como
“ESPECIE EN FORMACIÒN”, teniendo a los primeros homínidos como sus
infantes, a las versiones de Homo erectus como adolescentes y al sapiens
/ sapiens como adulto totalmente consumado, dentro y fuera de àfrica.
Cien años del Fraude del Hombre de Piltdown.
Por Mariano
Magnussen Saffer. Director
de Grupo Paleo e Integrante del Museo Municipal Punta Hermengo.
marianomagnussen@yahoo.com.ar Tomado de Magnussen Saffer,
Mariano (2012).
Cien años del Fraude del Hombre de Piltdown.
Paleo, Revista
Argentina de Paleontología. Boletín Paleontológico. Año 10.
76: 10-12.
En 1912 se presentó en la Sociedad Geológica de Londres una comunicación
sobre el hallazgo del “eslabón perdido” entre el hombre y los simios. El
descubrimiento del llamado Hombre de Piltdown produjo gran estruendo en
el mundo científico. Pero, en 1953, se “descubre” el fraude del
Eoanthropus, del Hombre de Piltdown. Algunos autores sospechan que el
famoso paleontólogo y poeta místico, Teilhard de Chardin, estuvo
involucrado en el fraude, que sabía que era falso pero que colaboró para
buscar notoriedad. ¿Inocente o culpable? Por Leandro Sequeiros.
El hombre de Piltdown
es conocido por ser uno de los más grandes fraudes en la historia de la
paleoantropología, principalmente porque se creyó verdadero durante
cuarenta y cinco años, desde que se anunciara su descubrimiento en 1908,
hasta 1953 cuando el fraude fue finalmente expuesto.
La historia de
este engaño comenzó y se basó en unos restos óseos (en concreto
un cráneo parcial, un diente suelto y una mandíbula con dientes)
descubiertos en Inglaterra en 1912, en Piltdown, un pueblo de
Sussex. Un obrero los encontró en una cantera, y se los entregó
al arqueólogo aficionado Charles Dawson, que los presentó, junto
con el eminente paleontólogo Smith Woodward (del Museo
Británico), en la Sociedad Geológica de Londres. Durante años,
se mantuvo el debate sobre el origen de estos restos, y la
prensa dijo que muy probablemente correspondieran al eslabón
perdido, denominándolo Eoanthropus dawsonii. Estos restos
fueron aceptados por la comunidad científica sin mayores
análisis, debido principalmente a que era perfecto e idéntico a
la idea de aquella época sobre el eslabón perdido.
La idea de esa época
era que el eslabón tenía que haber tenido un gran cerebro pero
igualmente presentar rasgos simiescos, y posteriormente haber
evolucionado a una apariencia humana; idea contraria a la existente
ahora y que presentan los fósiles verdaderos.
No
obstante, comenzaron a surgir cada vez más dudas sobre la antigüedad y
el origen de esos restos. Finalmente, el dentista A.T. Marston,
determinó que los dientes de ese esqueleto correspondían evidentemente a
un orangután, el diente suelto a un mono y el cráneo a un ser humano (Homo
sapiens): a partir de entonces, los análisis del contenido en flúor
de los huesos demostraron que el enterramiento había sido intrusivo, así
como que el color ferruginoso oscuro de los huesos se debía a un
tratamiento químico, para uniformar las diferencias de color entre la
mandíbula (más moderna) y el cráneo (más antiguo).
Nadie
sabe quién cometió el fraude, y algunos lo atribuyen a los descubridores
originales, señalando sobre todo a Dawson, motivado por el hecho de que
en las islas británicas no había sido descubierto ningún fósil humano,
mientras que en el resto de Europa y fundamentalmente en África sí. Sin
embargo, el profesor Douglas dejó a su muerte una cinta magnética en la
que señalaba que el autor de la falsificación fue el archifamoso
profesor Sollas, que pretendía con ello desprestigiar a su rival
Woodward. A pesar del fraude, se ha erigido, por suscripción popular, en
el lugar donde se descubrieron los huesos, un monumento honorífico a
estos restos: el propio Woodward asistió a la inauguración.
Igualmente,
existen teorías diversas que han atribuido la invención a
algunos de los hombres más famosos de la época, incluyendo a
Arthur Conan Doyle y a Teilhard de Chardin. Por mucho tiempo se
acusó a Dawson de ser el único culpable en el engaño, pero Gould
asegura que su investigación muestra que Teilhard, que acababa
de ser ordenado sacerdote y que en ese entonces estaba
estudiando paleontología, participó en la “conspiración de
Piltdown.” Gould dice que algunos de los huesos que se
encontraron en las fosas de Piltdown provenían de países en los
cuales Teilhard había recogido especímenes en viajes anteriores.
Además, en las cartas que Teilhard envió a uno de los
científicos que descubrieron el engaño, Gould afirma que
Teilhard mintió para ocultar su participación en la intriga.
"El hombre de Piltdown"
es el nombre con el que se conoce a cierta sección de la obra del
compositor británico Mike Oldfield llamada Tubular Bells,
compuesta en 1973. La sección "El Hombre de Piltdown" fue el resultado
de la petición que su discográfica (Virgin Records) le hizo con el fin
de añadir una parte vocal a Tubular Bells. En dicha parte, el
compositor citado distorsiona su voz para crear un timbre que puede
aludir a sonidos simiescos.
Bibliografia Sugerida.
A
Framework of Plausibility for an Anthropological Forgery: The Piltdown
Case, Michael Hammond, Anthropology, Vol 3, No. 1&2, May-December, 1979.
Di Trocchio, Federico (2007). Las mentiras de la ciencia.
Alianza Editorial. ISBN 978-84-206-3988-8.
Piltdown Man-The
Missing Links, L. Harrison Matthews, serie de artículos aparecidos en New
Scientist entre 30 abril de1981 y julio de 1981.
The
Panda's Thumb, Stephen Jay Gould, W.W.Norton and Company, New York, con
el ensayo "Piltdown Revisited".
The
Piltdown Papers, Frank Spencer, Oxford University Press, London 1990,
ISBN 0198585233, xii, 282 p. : ill. ; 25 cm.
Idas y vueltas de la Paleontología. Cuando un fósil cae
en desgracia.
Un fósil se vuelve
famoso con cientos de titulares alrededor del mundo, pero luego nadie se
acuerda de él. ¿Es que la paleontología sólo es generar nombres en latín
y descubrir fósiles? El curso normal de la ciencia no se refleja en los
medios, veamos qué es lo que sucede con un fósil mediático cuando se le
ha pasado su cuarto de hora.
El año pasado se dio a
conocer un fósil con bombos y platillos, como si fuese la presentación
de la última película de la estrella máxima del cine. Tal vez se
acuerden. Uno pequeño, precioso, con forma de monito, y que prometía
revolucionar la paleontología. Sus descubridores proclamaban que era el
famoso eslabón perdido. ¿Ahora se acordaron? ¿No? Es el Darwinius
massillae, apodado Ida.
No creo que nadie lo
recuerde por más famoso que se haya vuelto por unos días. Por lo general
los únicos fósiles que la gente suele tener bien presentes, como si se
tratase de las celebridades del mundo paleontológico, son el
Tiranosaurus rex y el mamut, seguramente gracias al cine. La
ciencia en general y la paleontología en particular funcionan de una
forma compleja que suele quedar fuera de las noticias, ya que el curso
normal de la ciencia no genera titulares. Sí se consiguen tapas y
titulares rimbombantes cuando hay descubrimientos. Pero claro, si uno
sólo se guiase por esto podría pensar que la paleontología consiste
únicamente en nombrar fósiles uno atrás del otro. A veces los
periodistas, tal vez entusiasmados por los científicos, solemos exagerar
esos descubrimientos diciendo que llegan a revolucionar la ciencia, para
que resulten más atractivos que poner simplemente un nombre en latín y
describir cómo es esa nueva criatura.
Sin bien esto
genera que uno se sorprenda ante el descubrimiento, pronto se
olvida el nombre del fósil. Así al poco tiempo la nueva criatura
cae en el olvido mediático, recluyéndose únicamente en la
literatura científica. Es que cuando una de estas criaturas es
mostrada como capaz de cambiar todo lo conocido hasta el
momento, no alcanza con presentarla. La verdadera ciencia viene
después. Cuando todos los científicos de la comunidad comienzan
a analizar y poner a prueba lo que han presentado los
descubridores.
Tomemos el caso de Ida,
el fósil que les mencionaba antes: Darwinius masillae. Se
lo presentó en sociedad en mayo de 2009. Se decía que era un fósil clave
para el estudio de la evolución humana y de los primates. Es una
criatura pequeña, similar a un lémur de la actualidad, pero con 47
millones de años de antigüedad. Se dijo que era el eslabón perdido, los
mismos descubridores lo dijeron en rueda de prensa, y así titularon el
documental que hicieron. Un fósil magnífico desde todo punto de vista,
ya que está en perfecto estado de conservación, y totalmente completo.
Sin hablar de que aporta información clave para conocer cómo eran los
primates de esa época, que por lo general son conocidos por huesos
aislados, y a veces un único diente. Darwinius había sido
descubierto en 1983 en una mina de carbón de Alemania por un
coleccionista privado, que se lo vendió por millones a Jorn Hurum en
2007. Claro, Hurum quería recuperar algo de ese dinero invertido, por lo
que organizó una presentación del fósil a todo trapo.
Lo llamaron “El
Enlace”, el eslabón perdido, el vínculo entre los antiguos primates y
los primeros ancestros humanos. Permitieron que los periodistas se
confundieran y lo llamasen el ancestro directo de monos y humanos,
cuando en realidad en el artículo científico que lo presentaba sólo se
decía que estaba cerca de nuestros ancestros en el árbol evolutivo. Para
que se ubiquen, nosotros y los monos modernos pertenecemos a una
división biológica llamada haplorrinos, luego hay otro tipo de primates
conocidos como strepsirrinos, dentro de los cuales están los lémures
actuales. Al Darwinius lo ubicaban entre esos dos grupos,
pero más cerca de nosotros. De este modo podría ayudar a comprender cómo
habría sido el antepasado común que nosotros compartimos con los monos
actuales.
Pero el show mediático
que generó Darwinius, con súper rueda de prensa, un libro
y documental incluido, desapareció pronto de los medios de comunicación,
y terminó cayendo en el olvido general. Los científicos no lo dejaron de
lado, para ellos el asunto recién comenzaba, ya que al poco tiempo
salieron artículos científicos que contradecían las conclusiones de los
descubridores. Decían que Darwinius era un muy buen fósil,
pero que no estaba en nada relacionado con nosotros o nuestros
ancestros, sino con los lémures. Sin embargo los descubridores Jorn
Hurum, Philip Gingerich y colegas, permanecieron en silencio hasta hace
poco, cuando respondieron a sus “agresores”. Aunque lo hicieron
repitiendo lo que ya habían dicho en una carta enviada al Journal of
Human Evolution.
Tras las
bambalinas de los medios masivos, en las publicaciones
científicas el debate giraba en derredor de si Darwinius
era un haplorrino o un strepsirrino. Esto no genera titulares,
pero para el mundo paleontológico es muy importante. Un lector
lego en el tema podrá preguntarse ¿y qué importa si pertenece a
uno u otro grupo?
Si fuese un haplorrino,
como sostienen los descubridores, entonces habría estado cerca de los
ancestros de los primates antropoides, o sea los monos y nosotros. Pero
si es un strepsirrino, como sostiene el resto de la comunidad
paleontológica, entonces habría pertenecido a un linaje extinto en otra
rama bastante alejada de nuestro árbol evolutivo. Siendo Darwinius
un fósil tan completo aporta infinitas posibilidades de estudio
y también hace que el debate parezca casi tonto, ya que no tendría que
haber mucha duda al estudiar a fondo cada uno de sus huesos. Su anatomía
debería casi hablar sola indicando a qué grupo pertenece.
Pero el debate surge
porque los dos grupos de científicos en discusión utilizan diferentes
métodos de comparación. Por ejemplo: en el artículo original en el que
se presentó a Darwinius, Hurum y colegas habían comparado
al fósil exclusivamente con primates vivos. Para los demás paleontólogos
esto parecía irrelevante, ya que las características elegidas se basaban
en los tejidos blandos, que obviamente en los fósiles no se conservan,
sólo quedan los huesos. De este modo pudieron encontrar muchas
características en común entre Darwinius y los monos actuales.
Pero desde principios
del siglo veinte que no se hace este tipo de comparación, ya que si es
un fósil se compara con otros fósiles cercanos en el tiempo, y con la
anatomía ósea de los animales actuales. Otros científicos compararon a
Darwinius con primates tanto fósiles como vivos. Uno de
ellos, Erik Seiffert y colegas, compararon 360 características
anatómicas con 117 primates diferentes. Y lo que descubrieron es lo que
acepta la mayoría hoy en día, que Darwinius está más
emparentado con la rama alejada de nosotros, la de los strepsirrinos,
los llamados adapiformes, que tenían forma de lémur. Sin embargo, los
descubridores de Darwinius publicaron un estudio, con
nuevas comparaciones. Pero otra vez dejaron fuera a los fósiles, e
incluso adujeron que era inútil compararlo con primates extinguidos.
Según ellos, como los haplorrinos y los strepsirrinos son grupos que se
establecieron en base a primates vivos, ellos debían comparar a
Darwinius con primates vivos para saber si pertenecían a uno u
otro grupo. No aportaron nada nuevo.
El problema es que al
no utilizar fósiles en la comparación se pierde la perspectiva.
Darwinius es un fósil de 47 millones años de antigüedad. Al ver
características de un fósil tan antiguo que son similares con las de
animales vivos actualmente, nos olvidamos de que en el medio hubo 47
millones de años de evolución, que es mucho tiempo. Lo que parecen
características compartidas podrían haber evolucionado dos veces en dos
linajes diferentes y en épocas diferentes. Al comparar a Darwinius
con otros fósiles se puede ver que los de nuestra línea evolutiva,
posteriores a Darwinius, no tienen esas características.
Esto indica que los
primates actuales no las obtuvieron de él, ni de un pariente cercano,
sino que ocurrió lo que se conoce como evolución convergente, que es
cuando una característica similar evoluciona en linajes diferentes y en
épocas diferentes. Darwinius habría pertenecido a una rama
que luego se extinguió. Lo interesante de este debate es que muestra
cómo funciona la ciencia, qué camino sigue luego de que en los medios
masivos se anuncia una nueva especie. Y éste es apenas uno de los
debates. Luego vendrán los que estudiarán qué comía, cómo era su
ambiente, qué costumbres tendría, etcétera. La paleontología no termina
en el anuncio, éste es apenas el inicio del proceso que llevará a los
científicos a comprender el pasado evolutivo. El debate y el trabajo que
lleva a un anuncio tienden a permanecer en la oscuridad. Pero sólo es
noticia si es nuevo e interesante. No nos olvidemos de que el ser humano
ansía lo novedoso, los chismes. Los entretelones no importan tanto.
La expansión de Homo Sapiens por Asia
meridional desde la perspectiva arqueológica.
Continuando con la
serie de expansión de Homo sapiens, en esta ocasión analizamos la
expansión de Homo Sapiens por Asia meridional desde el punto de vista
de la arqueología, lo cual es una tarea sencilla y complicada al mismo
tiempo, dependiendo hacia adonde miremos.
Si miramos atrás y
vemos la salida de nuestros antepasados de África atravesando la
península de Arabia y llegando hasta la India, podemos apreciar que la
ruta de acceso y el intervalo temporal no dejan muchas dudas al
respecto; así mismo, esta ruta presenta una clara continuidad hasta el
sudeste asiático y Australia avalada por el registro arqueológico.
La situación se
complica cuando situamos el punto de mira en la llegada de Homo Sapiens
a China, donde la evidencia arqueológica presenta grandes complicaciones
debido a recientes dataciones de restos fósiles de Homo Sapiens que
rondarían los 100.000 años de antigüedad, y por la dificultad para
encontrar elementos de transición del paleolítico inferior al
paleolítico medio en esta región
Volviendo al principio, según explicaba Maju en el post anterior de esta
serie, parece que la salida fuera de África se produjo por una doble vía
a través del corredor de Palestina y cruzando el estrecho de Bab al
Mandeb que separa África de Yemen, expandiéndose por la península de
Arabia favorecidos por el clima (Pluvial de Abassia) y siguiendo los
abundantes cursos fluviales entonces existentes en Arabia. El yacimiento
de Jebel Faya, parece darnos una pista sobre la continuidad de esta ruta
hasta la India, cruzando a través del estrecho de Ormuz.
Tradicionalmente algunos autores han tratado de explicar la
ruptura de esta ruta de expansión primigenia por el Próximo
Oriente aludiendo a la catástrofe de Toba, una súper erupción
volcánica que aconteció hace unos 75.000 años en Sumatra,
Indonesia, y que provocó un descenso de las temperaturas a nivel
mundial conduciendo a muchas poblaciones de seres humanos a un
cuello de botella evolutivo, a una casi extinción. Sin embargo,
esto no es suficiente para explicar por qué justo en ese momento
parece que la población de neandertales aumenta en el Próximo
Oriente.
En cualquier caso,
parece que ni la catástrofe de Toba ni la llegada de los neandertales al
Próximo oriente supuso un impedimento para la expansión de Homo Sapiens
hacia el sudeste asiático y Asia oriental, como veremos, ni la ruptura
de esa ruta de expansión iniciada en Skhul-Qafzeh, sino un cambio de
dirección hacia otras zonas.
En la India, las investigaciones de Michael Petraglia han puesto de
manifiesto la similitud de algunas industrias con tecnocomplejos propios
de la Middle Stone Age africana, avalando de esta forma la expansión de
Homo sapiens y su llegada a la India en torno a los 80.000 años
También se ha
evidenciado la existencia de tecnología lítica en algunos yacimientos
por encima y por debajo de la capa de ceniza de la erupción de Toba, lo
que demuestra que los seres humanos del paleolítico medio de India
sobrevivieron a esta tragedia; seguramente homo sapiens conviviera en la
zona con otros seres humanos extintos antes de la catástrofe, y puede
que esta tragedia afectara de distintas formas a los distintos grupos de
seres humanos; lo único que sabemos con certeza es que homo sapiens
sobrevivió y siguió su proceso de expansión.
La presencia de Homo
sapiens en India hace unos 80.000 años a día de hoy parece un hecho
incontestable desde el punto de vista arqueológico. Yacimientos como
Jwalapuram, con esta datación (80-78 ka bp aproximadamente) así lo
corroboran, donde según Petraglia hay una industria de afinidad
sudafricana. Otros yacimientos datados entre 85-74 ka bp como Samnapur o
los situados en Belan Valley reforarían la continuidad de esta ruta de
expansión de Homo Sapiens, bien siguiendo una ruta costera o bien
siguiendo el curso fluvial del río Narmada por el interior.
Pero a pesar
de la evidencia clara de esta ruta de expansión sapiens para el
intervalo temporal que comentábamos (85-74 ka bp), el
paleolítico medio de la India no está exento de interrogantes
difíciles de responder; el yacimiento de Didwana presenta
tecnologías "Modo 3" o de industrias sobre lascas en unas fechas
que oscilan entre los 150-100 ka bp. Si normalmente aceptamos
que las industrias conocidas como Modo 3 se adscriben bien a
neandertales bien a Homo sapiens, debemos asumir que este
yacimiento fue ocupado por una de estas dos especies de seres
humanos; el problema es que no parecen existir evidencias de
neandertales en esta zona, pero tampoco encajan bien las fechas
para una llegada tan temprana de sapiens a India (a no ser que,
como veremos, demos por válidas las dataciones de Homo sapiens
tempranos en China que podrían representar una continuidad de
esta yacimiento).
En Sri Lanka hay
evidencias de ocupación humana desde hace 125.000 años, en Bundala, con
industria lítica sobre cuarzo considerada de paleolítico medio, por lo
que tenemos otra duda acerca del tipo humano que las realizó.
Para resolver este enigma de la autoría de la tecnología de Didwana y
Bundala, podemos mirar al valle Narmada, donde hay restos fósiles de
seres humanos de difícil clasificación en Hathnora,
que bien podrían ser Homo Heidelbergensis con alguna afinidad hacia los
neandertales; los restos de estos seres humanos de Narmada están datados
en un rango temporal muy amplio, por lo que tampoco ayudan mucho a
resolver este enigma, y además están asociados a tecnología achelense.
En cualquier caso, Homo sapiens ocupa la India hace unos 80.000 años y
continua su viaje llegando hasta el sureste asiático y Asia oriental.
A partir de aquí, todo cuadraría perfectamente de no ser por las
recientes dataciones en la región de Guangxi-Zhuang, China, donde parece
ser que hay restos fósiles de Homo sapiens en Zhirendong con una
antigüedad de 110-100 ka bp, y en Liujiang, con una datación que oscila
entre los 139-68 ka bp. Sin embargo, no está muy claro que se puedan
atribuir estos restos de Zhirendong a humanos anatómicamente modernos,
se podría tratar de algún otro ser humano extinto cuyos márgenes de
variabilidad morfológicos puedan estar cerca de los humanos modernos.
Por el contario, los
restos de Liujiang parecen claramente de Homo sapiens, sin embargo las
dataciones son controvertidas. En cualquier caso la polémica está
servida, ya que las dataciones no cuadran con la llegada de Homo sapiens
a China, las fechas que se proponen son muy antiguas. Habrá que esperar
a que aparezcan más restos para que todo el mundo asuma su pertenencia a
Homo sapiens. En esta misma región hay restos de Homo sapiens en Laibin,
con unos 41.000 años de antigüedad.
La fecha de 68 ka bp
puede parecer más lógica; en cualquier caso, en China también hay restos
fósiles de Homo sapiens en Huanlong Cave, en la región de Hubei, datados
en unos 66.200 años, y en Tianyuan, datados
en 40.500 años.
Tras la
erupción de Toba,la expansión de Homo sapiens continua y se
incrementa el número de yacimientos arqueológicos por el sudeste
asiático; esta expansión seguramente se vio favorecida porque la
erupción de Toba redujo drásticamente las poblaciones de Homo
erectus, el tradicional habitante de esta zona. Las poblaciones
de sapiens avanzan y se encuentran con las últimas poblaciones
de erectus, muy mermadas en número tras la catástrofe.
En este mapa 2 los
yacimientos están representados en color azul para diferenciar el
intervalo temporal del primer mapa; un dato interesante es que mientras
la expansión de sapiens continúa hacia Asia meridional, observamos la
llegada de los neandertales al Próximo oriente (estrellas azules), y el
posible inicio de otra expansión sapiens representada en el niño de
Taramsa y en la aparición de algunos de la “cultura Emiriense”, que
quizás será la que se convierta con paso del tiempo en el ahmariense y
el auriñaciense que marcarán el inicio de una nueva expansión, esta vez
hacia Europa y Asia central; pero esta expansión será tratada en los
siguientes artículos.
En Laos tenemos
evidencias de restos fósiles de Homo sapiens en Cave of Monkeys con una
antigüedad entre 63.000 y 46.000 años.
Con una antigüedad
de 67.000 años tenemos en Callao Cave, Filipinas, restos fósiles que son
de dudosa atribución, ya que se trata de un metarso, y se podría asociar
no sólo a Homo sapiens, sino también a Homo erectus; así que no son
concluyentes de momento, pero en todo caso encajarían bien con estas
fechas que estamos viendo para la llegada de sapiens a esta zona.
En Borneo tenemos
restos fósiles de Homo sapiens en Niah Cave, datados en unos 40.000 años
de antigüedad.
En la isla de Flores
tenemos material arqueológico en Liang Bua datado entre 74-61 ka bp, que
seguramente puedan encajar a Homo sapiens, auque hay que recordar que en
esta zona es donde aparecieron los famosos restos del Homo Floresiensis
pero con dataciones posteriores (18 ka bp).
En Timor oriental,
en la cueva de Jerimalai, se encontraron
infinidad de utensilios de piedra (más de 9.700) y casi 40.000 huesos de
hasta 22 familias diferentes de pescados, además de fósiles de otros
animales terrestres del entorno, de los que también se alimentaron los
seres humanos., con una antigüedad que llegaría hasta los 42.000 años.
Algunos de los vestigios pertenecen a atunes, lo que implica que los
habitantes de aquel lugar pescaban desde embarcaciones en alta mar.
Parece ser que 15 de estas especies fueron explotadas en el periodo más
primitivo de ocupación de la cueva, hace entre 42.000 y 38.000 años. Hay
que poner en relación estos pueblos con toda seguridad con los primeros
colonizadores de Australia.
En el norte de Corea
tenemos restos de Homo sapiens en Ryonggok Cave, con unos
48.000 años de antigüedad, mientras que en el sur tenemos las evidencias
de Heungsu Cave, cuyas dataciones más antiguas rondan los 40.000 años.
La ocupación de Japón
ha estado envuelta en cierta polémica en los últimos años debido a una
serie de fraudes arqueológicos; al margen de polémicas parece que hay un
consenso generalizado en aceptar una ocupación más o menos clara en
torno a los 40.000 años; a partir de los 38 ka bp se produce un aumento
de la densidad de yacimientos en torno al lago Nojiri, con tecnología de
tipo paleolítico medio-superior.
Hay algún
artículo de prensa que habla de artefactos más antiguos,
llegando a proponer fechas entre los 120-70 ka bp, pero de
momento no hay nada concreto.
Parece más
coherente aceptar que tras la llegada a India hace unos 80.000
años, Homo sapiens continua con su expansión llegando al sudeste
asiático hace aproximadamente unos 70.000 años. Estas fechas
cuadran también con la subsiguiente colonización de Australia,
que se produjo hace entre 60.000 y 50.000. La colonización de
toda la parte de Asia oriental se produjo hace entre unos 68.000
años y unos 40.000 años.
No cuadran las
dataciones de Homo sapiens de 100.000 años en China, sin embargo no
podemos tampoco descartarlas ya que podrían representar la continuidad y
el éxito de una expansión de Homo sapiens temprana de las que quizás las
primeras evidencias fueran las de Skhul-Qafzeh.
La colonización de
Asia central parece que no tiene relación con esta primera expansión
iniciada hacia Asia meridional; parece que una nueva expansión iniciada
en el Próximo Oriente pudo llevar la tecnología auriñaciense a esta zona
y después se proyectaría a Europa, o quizás de Europa a Asia central.
Por lo tanto será
tratada con detalle en los siguientes artículos. No obstante adjunto
este mapa final sobre la colonización de Asia en distintos colores que
representan distintas fases de colonización.
Fuentes:
-
Out of
Africa: new hypotheses and evidence for the dispersal of Homo sapiens
along the Indian Ocean rim
-
New
geochronological, paleoclimatological, and archaeological data from the
Narmada Valley hominin locality, central India
- Prehistoria I. Las primeras etapas de la Humanidad. Petraglia, M., Haslam, M., Fuller, D., Boivin, N., &
Clarkson, C. (2010). Out of Africa: new hypotheses and evidence for the
dispersal of along the Indian Ocean rim Annals of Human Biology, 37
(3), 288-311.