|
Los Xenarthros
Pilosa o
Perezosos Gigantes,
en el registro fosilífero
local.
Por
Mariano Magnussen. Laboratorio Paleontológico del Museo
de Ciencias Naturales de Miramar. Fundación Azara. Laboratorio de
Anatomía Comparada y Evolución de los
Vertebrados.
marianomagnussen@yahoo.com.ar
. Ilustraciones Daniel Boh
La historia
evolutiva de los Xenarthros, edentados
sudamericanos, o también conocidos como
perezosos, es extensa e increíble. Algunas
criaturas eran muy pequeñas y simples (perezas,
perezosos de dos dedos u osos hormigueros), a
gigantes bestias del pasado (megaterios y
lestodontes), incluyendo a aquellos de piel
acorazada (gliptodontes y armadillos), único
entre todos los mamíferos extintos y vivientes
en todo el mundo. El único carácter externo que
tienen en común entre ellos, son las uñas en
formas de garra grandes y robustas, adaptadas
para funciones diversas, tanto en las formas
terrestres (excavar) como en las arborícolas
(agarrarse a las ramas).
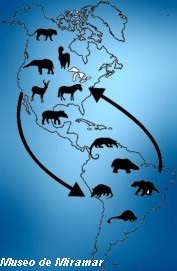
Los Xenarthros,
están representados por los del Orden Pilosa.
Casi todos los taxones de este grupo de
mamíferos se registran en América del Sur desde
el Paleoceno tardío, hace 56 millones de años,
hasta la actualidad. También existen varios
registros en la Antártida que pueden ser
considerados dentro del grupo de los Xenarthros
en la época del Eoceno, hace unos 40 millones de
años antes del presente, y algunos pocos en
Norteamérica, que llegaron allí hace 2 millones
de años, durante el Gran Intercambio Biótico
Americano, luego de la unión de ambas Américas,
generando asi, un intercambio faunístico.
 |
En nuestra área, el
registro más antiguo (por la juventud de los
sedimentos) corresponden al Plioceno medio, unos
4 millones de años. El Museo de Ciencias
Naturales de Miramar, ha recuperado fósiles de
Proscelidodon, Glossotheridium y
Scelidotheridium, antecesores de
aquellos de mayor tamaño que vivieron durante el
Pleistoceno. Estos eran mas pequeños (pero no
tanto) de las formas posteriores
Así mismo, en estos
sedimentos Terciarios, hemos observado
crotovinas, es decir, antiguas madrigueras
rellenas de sedimentos que de depositaron en su
interior con un diámetro superior al metro y
medio, lo que coincidiría a estas formas de
Pilosas, cuyas estructuras son mejores conocidas
y estudiadas durante el Pleistoceno regional.
|
Además, en 2021, un
equipo conformado por la Fundación Azara, el
Laboratorio de Anatomía Comparada y Evolución de
los Vertebrados y del Museo miramarense,
presentaron el hallazgo de un cráneo juvenil del
Megaterio (Megatherium sp) más antiguo de
Argentina, encontrado en las proximidades de
Miramar en sedimentos del Plioceno, que, junto a
otros restos de Bolivia, son los más antiguos
del mundo para el presente género. Sin dudas se
trata de una versión anterior al gigante del
Pleistoceno, pero al tratarse de un individuo
juvenil, no se le pudo nombrar como nueva
especie.
Durante el
Pleistoceno, entre 2,5 millones de años a 11 mil
años antes del presente, el orden Pilosa, tiene
una gran variedad de tamaños y formas. Se
caracterizan por sus dientes simples sin
esmalte (como el marfil), para triturar plantas
semiduras y fibrosas, y en algunas especies,
presentaban dientes modificados con forma de
caninoformes (colmillos) en su extremo frontal,
largas lenguas prensiles, articulaciones
adicionales en las vértebras, huesos largos
robustos y miembros dirigidos hacia adentro,
estructura osteodermica, es decir, huesecillos
esféricos y redondeados separados entre sí,
formando un escudete defensivo imperceptible en
la piel, además, de garras enormes para realizar
madrigueras para refugio o protección de
juveniles, búsqueda de raíces y tubérculos, y
control en la temperatura corporal.
 |
Entre sus
representantes, que conformaron la Megafauna, es
decir, animales que superaban la tonelada de
peso, encontramos en su máximo exponente, el
Megatherium americanum, el más grande
de todos los mamíferos terrestres de la Era
Cuaternaria. Superaba los 4,5 metros de altura
cuando se paraba sobre sus patas y con un peso
estimado de esta especie es de 4 toneladas. Si
bien era herbívoro, nuevos hallazgos muestran
que consumían proteínas de origen animal, los
cuales cazaba, o tal vez, por su tamaño, robaba
presas a otros mamíferos depredadores, o se
alimentó ocasionalmente de carroña. |
En 2019, nuestro
Museo local, junto a colegas del Lacev (Macn -
Conicet) y Fundación Azara, presentamos nuevos
restos de Megatherium filholi, una
especie de Megaterio enana. Los fósiles nos
muestran, tal como sospechó el Perito Moreno,
quien descubrió los primeros restos de esta
especie en 1888, que el Megatherium
filholi era un enano entre los gigantes,
porque, pese a que medía unos tres metros de
largo y pesaba poco más de una tonelada, es muy
pequeño al ser comparado con su pariente más
cercano, el Megatherium americanum.
Desde entonces se sospechaba de la validez de
esta especie, que fue ratificada por los
hallazgos de sus restos en las proximidades de
Miramar.
Otro de los gigantes
con mayor registro en Miramar, fue
Lestodon arnatus, es el segundo perezoso
de mayor tamaño. Aunque resientes restos fósiles
hallados cerca del muelle de pescadores de
nuestra ciudad, demuestra la existencia de
grandes individuos, con talla similar o tal vez
superior al anterior. El Lestodonte, al igual
que sus parientes era un herbívoro, el cual
recogía su comida a orillas de los pantanos o
cauces fluviales. Median aproximadamente unos 4
metros de longitud. Su cráneo tenía una región
rostral muy ensanchada. Los huesos de sus
miembros eran grandes y gruesos. Su dentición
era simple, pero resaltaban hacia afuera de la
boca unos caniformes muy grandes, lo que se
convertía en una defensa contra los posibles
depredadores. Su masa estimada es de 2,5
toneladas.
 |
Otro Piloso gigante
fue Catonyx tarijensis, un Pilosa
de hábitos terrestres, corpulento, con cabeza
grande y una cola larga y pesada. Los pies
largos y con garras envueltas hacia adentro al
igual que los otros Pilosa ya descriptos, por lo
tanto, caminaba apoyando sus nudillos. Es
probable que pudiera ponerse en dos patas y
mantener el equilibrio con la cola, utilizando
sus garras para llevarse el alimento a la boca.
Tenía 3.50 metros de largo y 1,70 de alto. Su
masa estimada fue de una tonelada. Es sumamente
escaso en toda Argentina, y más frecuente en el
centro – norte de Sudamérica. |
El
Scelidotherium leptocephalum, era de
tamaño menor al resto de los perezosos, pero no
dejaba de ser un gigante. Sin dudas, fueron los
mamíferos más comunes del paisaje en el
Pleistoceno miramarense, ya que sus restos
fósiles son muy abundantes en los depósitos
sedimentarios de esta edad geológica. Tenía 1,70
metros de alto y 3,50 de largo. Su peso estimado
es de una tonelada. El cráneo es alargado, bajo
y estrecho, con un rostro proyectado hacia
adelante. Su alimentación consistía de vegetales
que hallaba en áreas secas.
En el año 2002 se
halló en Miramar una paleocueva con galerías,
dos cámaras y garras marcadas en paredes y
techos. Su extensión superaba los 26 metros de
largo y un diámetro que llegaba a los 1,90
metros, y se podía observar su antigua conexión
con la superficie. En esta, se pudo recuperar
marcas de garras pares que coinciden con la
anatomía de Scelidotherium. Así
mismo, en 2004, personal de este Museo de
Ciencias Naturales de Miramar, recupero en la
localidad de Mar del Sud (15 kilómetros de
Miramar) el esqueleto de un adulto, abrazando a
un juvenil. La hipótesis, es que se trata de
una hembra junto a su cría, que murieron en su
madriguera hace más de 100 mil años antes del
presente, siendo al día de hoy, un registro
único. En este museo se conservan decenas de
restos de Celidoterio, mientras que en
exhibición se pueden observar un cráneo, una
mano completa, y un esqueleto completo.
 |
Mientras tanto,
Mylodon darwinii, es otro de los
grandes perezosos registrado en la zona, menor
que Megaterio y Lestodonte. Se alimentaba de
vegetales. Se calcula que medía aproximadamente
2,50 metros a la cruz, y pesaba unas
2,5 toneladas. Un singular hallazgo, fue
realizado a fines del siglo pasado. Proviene de
una caverna en el Seno de la Ultima Esperanza,
llamada también como "Cueva del Mylodon"
donde fueron encontrados excrementos del
animal y huesos de gran tamaño
diseminados en sedimentos que incluían
restos de carbón. |
Lo asombroso de
este descubrimiento, es que la pieza única
permitió conocer la estructura del cuero de
estos animales, que en su interior alojaba
pequeños huesecillos subesfericos (osteodermos)
que, si bien permitían flexibilidad a la piel,
convertían al Milodonte en un verdadero
acorazado imperceptible. Esta característica se
repitió en otros edentados, pero tal vez, con
menos densidad.
El
Glossotherium robustum, poseía gran
tamaño y hábitos terrestres, corpulento, con
cabeza grande y una cola larga y pesada. Tenía
3.50 metros de largo y 1,70 de alto. Su masa
estimada fue de una tonelada y su extinción
ocurrió durante el Holoceno medio, hace 8 mil
años antes del presente. Su cuerpo estaba
cubierto por una espesa y densa pelambre y
embebidos en la piel se encontraba un gran
número de huesillos (osteodermos), en forma y
tamaño variables, generalmente superiores a un
centímetro de diámetro. Su hábito alimenticio
era variado, constituido de hojas de graminias,
arbustos, árboles y posiblemente carroña. Hace
poco tiempo, personal del Museo de Ciencias
Naturales de Miramar y Fundación Azara, recupero
la pata trasera de un ejemplar joven (pelvis,
fémur, tibia etc) recubierta de centenares de
huesesillos dérmicos, lo que significa que la
pata estaba cubierta por el cuero al quedar
sepultada.
 |
Otra especie, con
menor frecuencia de hallazgos, la conforma
Scelidodon copei, el cual, posee un
cráneo pequeño, alargado, estrecho y bajo; con
las mandíbulas alargadas, pero con los
premaxilares más cortos. Sus dientes, en la
superficie masticatoria, son elípticos o
levemente triangulares, con la serie dentada
paralela y continua, siendo tanto las superiores
como las inferiores casi del mismo tamaño, lo
que denota su dieta herbívora, consumiendo
arbustos, hojas y raíces.
Su cuerpo era masivo y
probablemente cubierto de un grueso pelaje.
Midió unos 2.50 metros de largo por 1 metro de
altura, con un peso cercano a la tonelada. Sus
extremidades posteriores son largas y robustas,
las anteriores son más cortas, terminando ambas
en fuertes garras curvadas hacia el interior,
por lo que se desplazaba sobre el dorso de sus
manos y pies. |
El estudio de estos
ejemplares permite comprobar que durante el
Pleistoceno existieron marcadas variaciones en
la composición y distribución de la fauna
pampeana en general, y de los Xenarthros en
particular, hecho íntimamente relacionado al
Gran Intercambio Biótico Americano y a las
variaciones climáticas producidas durante el
Pleistoceno, sumado a un declive y
empobrecimiento ambiental. A comienzos del
Holoceno, los gigantes perezosos se habían
extinto para siempre.
Bibliografía
sugerida.
Alberdi, G. Leone y
E.P. Tonni (eds)- Evolución biológica y
climática de la región pampeana durante los
últimos cinco millones de años. Un ensayo de
correlación con el mediterráneo occidental.
Monografía del Museo Nac. Cs. Nat. Consejo
superior de investigaciones científicas. Madrid.
pp. 77-104.
Bonaparte José.
2014. El origen de los mamíferos. Fundación de
historia natural Felix Azara.. ISBN
978-987-29251-8-5.
Borrero L. 2009. La
evidencia evasiva: el registro arqueológico de
la megafauna extinta sudamericana. En: Haynes G.
(Ed.), Extinciones megafaunales americanas al
final del pleistoceno: 145–68. Springer Science,
Dordrecht.
Esteban, G.1996.
Revisión de los Mylodontinae cuaternarios (Edentata,
Tardigrada) de Argentina, Bolivia y Uruguay.
Sistemática, Filogenia, Paleobiología y
Paleozoogeografía y Paleoecología.
Esteban, G.1996.
Revisión de los Mylodontinae cuaternarios (Edentata,
Tardigrada) de Argentina, Bolivia y Uruguay.
Sistemática, Filogenia, Paleobiología y
Paleozoogeografía y Paleoecología. Tesis
Doctoral, Instituto Miguel Lillo, Facultad de
Ciencias Naturales, 235 p. Tucumán.
Fidalgo, F. y Tonni,
E.P. 1983. Geología y paleontología de los
sedimentos encausados del pleistoceno tardío y
holoceno de Punta Hermengo y arroyo Las
Brusquitas (Partido de General Alvarado y
General Pueyrredon, Provincia de Buenos Aires).
Ameghiniana 20 (3-4): 281-296.
Giacchino, A.,
Bogan, S., Boh, D., Magnussen, M. y Meluso, J.
M. 2020. La creación del Museo de Ciencias
Naturales de Miramar “Punta Hermengo” y sus
antecedentes (General Alvarado, provincia de
Buenos Aires, República Argentina). Historia
Natural (3ra serie), 9
Graham RW y
Lundelius EL 1984. Desequilibrio coevolucionario
y extinciones del Pleistoceno, en: Martin P.
Klein y RG (eds.), Extinciones cuaternarias: una
revolución prehistórica, pp. 223–249; Prensa de
la Universidad de Arizona (Tucson).
Magnussen Saffer, M.
2005. La Gran Extinción del Pleistoceno.
Boletín de divulgación Científico Técnico.
Museo Municipal de Ciencias Naturales Punta
Hermengo de Miramar, Prov. Buenos Aires,
Argentina. Publicación 3: pp 3 – 10 (Sec.Pal).
Magnussen Saffer,
Mariano. 2015. Los Mamíferos Xenartros más
característicos del Plioceno de la República
Argentina. Paleo Revista Argentina de
Paleontología. Boletín Paleontológico. Año XIII.
129: 12-13.
Nicolás R. Chimento,
Federico L. Agnolin, Diego Brandoni, Daniel Boh,
Mariano Magnussen, Francisco De Cianni y
Federico Isla (2020). A new record of
Megatherium (Folivora, Megatheriidae) in the
late Pliocene of the Pampean region (Argentina).
Journal of South American Earth Sciences.
Pascual, R. y O. E.
Odreman Rivas. 1971. Evolución de las
comunidades de los vertebrados del Terciario
argentino. Los aspectos paleozoogeográficos y
paleoclimáticos relacionados. Ameghiniana 8:
372-412.
Quintana, C. A.
2008. Los fósiles de Mar del Plata. Un viaje al
pasado de nuestra región. Buenos Aires,
Argentina. Fundación de Historia Natural “Félix
de Azara”. 242 pp.
Reig, O. A. 1958.
Notas para una actualización del conocimiento de
la fauna de la Formación Chapadmalal. I. Lista faunística
preliminar. Acta Geol. Lilloana 2:241-253.
Scillato-Yané, G.J.;
Carlini, A.A.; Vizcaíno, S.F.; Ortíz Jaureguizar,
E. 1995. Los Xenarthros. In Evolución biológica
y climática de la región pampeana durante los
últimos cinco millones de años. Un ensayo de
correlación con el Mediterráneo occidental
(Alberdi, M.T.; Leone, G.; Tonni, E.P.;
editores). Museo de Ciencias Naturales, Consejo
de Investigaciones, Monografías, p. 183-209.
Madrid.
Sebastian Apesteguia
y Roberto Ares. 2010. Vida en evolución: la
historia natural vista desde Sudamérica. Ed.
Vázquez Mazzini, 382 pp.
Simpson, G.G.
(1940). Review of the mammal-bearing tertiary of
South America. Proceedings of the American
Philosophical Society, 83: 649-710. |
Gliptodontes y Armadillos.
Los gigantes acorazados
hallados en la región.
Por
Mariano Magnussen. Laboratorio Paleontológico del Museo
de Ciencias Naturales de Miramar. Fundación Azara. Laboratorio de
Anatomía Comparada y Evolución de los
Vertebrados.
marianomagnussen@yahoo.com.ar
. Ilustraciones Daniel Boh
El aislamiento prolongado de
América del Sur, durante casi toda la Era Terciaria, es
decir, entre 60 y 3 millones de años antes del presente,
genero el desarrollo de una fauna muy particular, entre
ellos, los mamíferos del Superorden de los Xenarthros,
conformado por tres tipos morfológicos básicos, como los
osos hormigueros, de cuerpo alargado y hocico muy
prolongado, pelaje denso y costumbres terrestres o
arborícolas, seguido por los perezosos, de cuerpo
compacto, hocico corto, pelaje denso, de andar lento y
arborícolas, y por último, los armadillos, con el cuerpo
ancho y deprimido revestido dorsalmente con una coraza,
con poco pelo y de hábitos terrestres.
Justamente, estos últimos,
son los armadillos o perezosos acorazados, los edentados
del Orden Cingulata. Estos son muy frecuentes y con una
diversidad asombrosa localmente, tanto en Miramar como
en gran parte de la costa bonaerense, desde el Plioceno
hasta hace unos milenios, caracterizados por la
presencia de coraza dorsal, las cuales pueden tener
bandas móviles como los armadillos actuales o totalmente
rígida como los extintos gliptodontes (al menos en
estado fósil, ya que seguramente en vida, tuvo leves
movimientos) formados por osteodermos anexados, además
de poseer vértebras del cuello y del tronco fundidas,
incluso adheridas a la coraza. En el Pleistoceno, estos
verdaderos mamíferos acorazados, alcanzaron enormes
tamaños y variedades.
 |
Lo interesante de la
superficie expuesta de los osteodermos (placas
del caparazón), las cuales son de origen
dérmico, poseen una serie de figuras
ornamentales, que varían en todas las especies,
en sus distintos momentos de crecimiento, o
también en la ubicación de su cuerpo, lo que
facilita a los paleontólogos en su
clasificación. El más característico, es el
osteodermo del genero Glyptodon sp,
con forma de “flor”. |
En general, todas las
especies de gliptodontes se caracterizan por un cráneo
corto y ancho, también protegido con un escudete óseo, y
arcos cigomáticos bien marcado y desarrollado, para
adherir la musculación utilizadas para masticar. Todo el
cuerpo cubierto por un grueso caparazón, cuyo espesor en
algunos casos superaban los ocho centímetros. Los huesos
de la cadera no están soldados como en el resto de las
especies de los mamíferos, posiblemente, esta adaptación
posibilitaba que los gliptodontes pudieran pararse sobre
sus patas traseras, que eran cortas y muy gruesas,
utilizando la cola para apoyarse, las cuales estaban
recubiertas de unos anillos con púas óseas, o en otros
casos terminaban con un tubo óseo con púas corneas, lo
que les posibilitaba, vigilar el territorio, aparearse,
e incluso, dar a luz.
Su origen de los Cingulados
se remonta al Paleoceno, hace 55 millones de años, para
cuando ya estaban bien diferenciados, pero recién al
final del Plioceno y durante todo el Pleistoceno
tuvieron una gran diversidad, formas y tamaños.
Los Gliptodontes, se
originaron más tarde, durante el Eoceno, es decir, unos
45 millones de años antes del presente, encontrándose
sus restos en sedimentos del Terciario de la Patagonia y
Norte Argentino. Los rasgos fundamentales de esta
familia, es la presencia de una coraza sin bandas
móviles, colas protegidas con anillos móviles en forma
decrecientes, y algunos presentaban un tubo caudal óseo
con púas corneas en forma de defensa o ritual sexual, y
un casquete cefálico que protegía la cabeza. También
tenían la columna fusionada y soldada en gran parte al
caparazón dorsal, miembros cortos, pero gruesos y
fuertes.
 |
La gran cantidad de
especies encontradas desde el Plioceno regional,
muestran gran variedad, que citaremos a
continuación, las cuales, son las especies más
características de esta familia extinta, y tal
vez, la más popular entre los mamíferos fósiles
sudamericanos. En el Pleistoceno, los
Gliptodontes alcanzaron su máximo desarrollo,
tanto en variedad como en formas.
|
Al igual que otros
integrantes de la megafauna Sudamericana, se extinguen
sin dejar representantes vivientes. El deterioro
climático y ambiental, la llegada de nuevos predadores,
incluyendo los humanos, nuevos virus, fueron algunos de
los factores que determinaron para siempre la suerte de
los gliptodontes.
El Doedicurus
tuberculatus, fue la especie de mayor tamaño
dentro de la familia. Tenía 4,30 metros de largo y 2
metros de alto. La coraza tiene una forma muy
particular, presentando una especie de joroba en su
parte más alta, con placas más laxamente unidas que en
otros parientes de la época. Su cola estaba formada por
cuatro anillos, un tubo cilíndrico de un metro de largo
y en su término poseía una masa ósea con púas corneas,
convirtiéndose en una técnica de defensa contra los
habituales depredadores. Su masa estimada en vida es de
2 toneladas.
El Glyptodon muñizi, tenía
3,50 metros de largo y 1,4 de alto. Sus placas
engrosadas entre si varían de 1 a 7 centímetros de
diámetro, las cuales son fáciles de identificar por su
forma de "flor". Su cola carecía del anquilosamiento de
los anillos terminales en forma de tubo descriptos en
los otros géneros. Esta se encuentra formada por una
serie de anillos móviles de diámetro decreciente,
conformada por unos siete anillos con placas de forma
relativamente grotesca y en punta. De esta especie, se
exhibe un esqueleto muy completo.
 |
Por otro lado,
Panochthus tuberculatus, fue otro
gliptodonte de gran tamaño. Si bien, hemos
encontrado numerosas piezas de este enorme
animal, solo exhibimos en el parque del Museo,
una recreación en vida en tamaño natural.
Se
diferencian de los grandes y mediados
gliptodontes Cuaternarios por su coraza en forma
de casco prusiano.
Las
placas que forman este formidable caparazón son
gruesas, presentando un relieve con pequeñas
figuras circulares y uniformes finamente
punteadas. Su cola estaba constituida por
cuatro anillos móviles articulados, seguido de
un tubo con
numerosas púas de distinto tamaño. |
El Neosclerocalytus ornatus,
fue la más pequeña de todas las especies de los
Gliptodontes, que habitaron en América del Sur, la cual
alcanzaba apenas los 2,50 metros de largo y pudo pesar
unos 300 kilogramos. En las colecciones del Museo de
Ciencias Naturales de Miramar resguardamos una amplia
variedad de restos, e incluso una especie única,
mientras se exhiben un cráneo completo y la cola de esta
peculiar especie.
En el caso de los armadillos
con bandas móviles, estuvieron bien diversificados en
formas y tamaño, pero solo sobrevivieron dos especies
que habitan en el S.E bonaerense. Hemos registrado en
bosques y zonas abiertas de Miramar al peludo pampeano (Chaetophractus
villosus) y la mulita pampeana (Dasypus
hybridus). Debemos reconocer que, en otras
regiones del país y de América, aún hay una importante
variedad de armadillos, muy diferentes a los grandes
armadillos del Plioceno y Pleistoceno, pero
significativas para la mastozoología, aportando datos
del pasado biológico de estos animales.
 |
En una de las
vitrinas de esta sala, se exhiben los
Dasipodidos o armadillos, es una familia que
agrupa a animales cuyos cuerpos se hallan
protegidos por una coraza ósea y placas
dérmicas, la que se divide en dos partes, una la
coraza escapular y la otra la coraza pélvica,
separadas por varias bandas móviles que varían
en su cantidad según la especie. En la
actualidad se encuentran representados por
mamíferos de pequeño tamaño, como los Peludos y
las Mulitas, o el Tatú Carreta, como máximo
exponente en el norte argentino.
|
Entre las formas más comunes
del Plioceno, encontramos a Ringueletia simpsoni,
un armadillo de gran tamaño, de dieta variada, y que
construía sus propias cuevas, el cual vivió junto a
otros armadillos, como Paraeuphractus, Doellotatus, Macrochorobates, Chorobates, Tolypeutes,
Zaedyus, Holmesina, Kraglievichia, Scirrotherium,
Plaina, entre otros.
Durante la era Cuaternario
vivió un mulita gigante, llamada Pampatherium
typum, cuyo nombre significa "bestia de la
pampa". Es probable que su peso fuese de unos 225
kilogramos y superaba los 2,50 metros de longitud.
Construían grandes galerías subterráneas, con cámaras
amplias para proteger sus crías, las cuales, han sido
encontradas en los barrancos del Bosque del Vivero
Dunicola. En el Museo de Ciencias Naturales de Miramar
posee un ejemplar con varios restos de su esqueleto, y
otros ejemplares aislados.
Otro gigante extinto con
bandas móviles en su caparazón, fue Eutatus
seguini. Su registro fosilífero abarca desde el
Pleistoceno inferior (2 millones de años) hasta el
Holoceno temprano (8 mil años), encontrándose en algunos
casos, asociados a restos óseos de otros animales por
actividad antrópica, es decir, que han presentado
manipulación por antiguos grupos humanos que vieron en
esta zona a fines del Pleistoceno, en cuevas del sistema
serrano. Su coraza dorsal es robusta, y poseía unas 33
bandas móviles que cubrían las dos terceras partes del
caparazón. En cada mandíbula poseía de 9 a 10 dientes,
los cuales utilizaba para comer carne en descomposición,
huevos, larvas, caracoles y algunos tallos.
 |
El Propraopus,
era muy similar a los actuales Tatú carreta (Priodontes
maximus) que vive en el norte argentino,
pero su aspecto sería más parecido a la mulita
pampeana grande (Dasypus hybridus).
De alimentación fue
muy variada, compuesta principalmente por
gusanos, huevos, vegetales y animales en estado
de descomposición. La coraza sola, tiene unos 70
centímetros de largo, más la cola unos 40
centímetros. |
Otro representante del
Pleistoceno fue, Chaetophractus sp, un
armadillo fósil emparentado con el mismo género viviente
(el peludo pampeano). Restos fósiles de este y otros
géneros morfológicamente similares son hallados con
frecuencia en los afloramientos sedimentarios
correspondientes al Plioceno y Pleistoceno de Miramar,
esto quiere decir, que nuestro Peludo, ya era un
habitante frecuente en nuestra área en los últimos 3
millones de años.
A pesar de su
diversificación durante el pasado-reciente, hoy en día,
en nuestra región, quedaron reducidos a ejemplares que
no superan los 45 centímetros de largo y los 2
kilogramos de peso. Tanto el peludo como la mulita, son
omnívoros, incluyendo carroña a sus dietas, y construyen
sus madrigueras y refugios en campos, bosques, llanuras
o al costado de los caminos.
Bibliografia consultada.
Ameghino, F. 1889.
Contribución al conocimiento de los mamíferos fósiles de
la República Argentina. Academia Nacional de Ciencias,
1-1027.
De los Reyes, L.M;
Cenizo, M.M.; Agnolin, F.; Lucero, S.; Bogan, S.;
Lucero, R.; Pardiñas, U.F.J.; Prevosti, F. & Scanferla,
A. 2006a. Aspectos paleofaunísticos y estratigráficos
preliminares de las sucesiones plio-pleistocénicas de la
localidad Centinela del Mar, provincia de Buenos Aires,
Argentina. 90 Congreso Argentino de Paleontología y
Bioestratigrafía, 105.
Fariña RA, Vizcaíno SF.
1996. Hábitos locomotores del armadillo pleistoceno
Propraopus grandis (Mammalia, Dasypodidae): una
comparación
Fariña RA, Vizcaíno SF y
Bargo MS 1998. Estimaciones de masa corporal en
megafauna de mamíferos de Lujania (Pleistoceno
tardío-Holoceno temprano de Sudamérica). Mastozoología
Neotropical 5: 87-108.
Giacchino, Adrián y
Bogan Sergio. 2012. Colecciones. Colecciones Naturales y
antropológicas. Fundación de historia natural Felix
Azara. ISBN 978-987-27785-1-4.
M. T. Alberdi, G. Leona
y E. P. Tonni (editores). Evolución biológica y
climática de la región pampeana durante los últimos
cinco millones de años. Un ensayo de correlación con el
Mediterráneo occidental. Madrid, Museo Nacional de
Ciencias de Madrid, Consejo Superior de Investigaciones
Científicas, 1995, Monografías 12.
Magnussen Saffer, M.
2005. La Gran Extinción del Pleistoceno. Boletín de
divulgación Científico Técnico. Museo Municipal de
Ciencias Naturales Punta Hermengo de Miramar, Prov.
Buenos Aires, Argentina. Publicación 3: pp 3 – 10 (Sec.Pal).
Magnussen Saffer,
Mariano. 2015. Los Mamíferos Xenartros más
característicos del Plioceno de la República Argentina.
Paleo Revista Argentina de Paleontología. Boletín
Paleontológico. Año XIII. 129: 12-13.
Novas Fernando 2006.
Buenos Aires hace un millón de años. Editorial Siglo XXI,
Ciencia que Ladra. Serie Mayor.
Pascual, R. y O. E. Odreman Rivas.
1971. Evolución de las comunidades de los vertebrados
del Terciario argentino. Los aspectos
paleozoogeográficos y paleoclimáticos relacionados.
Ameghiniana 8: 372-412.
Pascual R., Ortega
Hinojosa D., Gondar y Tonni E. 1965. Las edades del
Cenozoico mamífero de la Argentina, con especial
atención a las aquéllas del territorio
bonaerense. Anales Comisión Investigaciones Buenos Aires
6: 165–193.
Quintana, C. A. 2008.
Los fósiles de Mar del Plata. Un viaje al pasado de
nuestra región. Buenos Aires, Argentina. Fundación de
Historia Natural “Félix de Azara”. 242 pp.
Scillato Yané, G.J.
1980a. Catálogo de los Dasypodidae fósiles (Mammalia,
Edentata) de la República Argentina. 2º Congreso
Argentino de Paleontología y Bioestratigrafía y 1º
Congreso Latinoamericano de Paleontología (Buenos
Aires), Actas 3: 7-36.
Sebastian Apesteguia y
Roberto Ares. 2010. Vida en evolución: la historia
natural vista desde Sudamérica. Ed. Vázquez Mazzini, 382
pp.
Vizcaíno, S.F., Milne,
N. and Bargo, M.S. 2003. Limb reconstruction of Eutatus
seguini (Mammalia, Dasypodidae). Paleobiological
implications. Ameghiniana 40(1): 89-101.
Tonni, E.P.; Fidalgo, F.
1979. Consideraciones sobre los cambios climáticos
durante el Pleistoceno tardío-Reciente en la Provincia
de Buenos Aires. Aspectos Ecológicos y Zoogeográficos
Relacionados. Ameghiniana, Vol. 15, Nos. 1-2, p.
235-253.
Tonni E. P., 2005. El
último medio siglo en el estudio de los vertebrados
fósiles. Asociación Paleontológica Argentina.
Publicación Especial 10, 50º Aniversario: 73-85.
Tonni E. P., 2016. Los
acantilados de la costa atlántica bonaerense y su
contribución al conocimiento geológico y paleontológico.
En: J. Athor y C. E. Celsi (eds.): La costa atlántica de
Buenos Aires. Naturaleza y patrimonio cultural.
Fundación de Historia Natural Félix de Azara - Vázquez
Mazzini Editores, pp. 42-65, Buenos Aires.
Los roedores fósiles
histricomorfos,
dregistrados en General
Alvarado.
Por
Mariano Magnussen. Laboratorio Paleontológico del Museo
de Ciencias Naturales de Miramar. Fundación Azara. Laboratorio de
Anatomía Comparada y Evolución de los
Vertebrados.
marianomagnussen@yahoo.com.ar
. Ilustraciones Daniel Boh
Cuando pensamos en el
término roedor, se nos viene a la mente las ratas y
lauchas. Pero en el pasado biológico como en la
actualidad, son algo más diversificados de lo que
creemos. Si bien no son nativos de Sudamérica, llegaron
en distintas oleadas migratorias, y se adaptaron
rápidamente al ambiente. Haciendo que varias formas
nuevas se conviertan en endémicas, es decir, que solo se
las encuentra en este lugar.
Entre algunos de los
roedores, encontramos los histricomorfos, es decir,
mamíferos placentarios con dos incisivos, de gran tamaño
de crecimiento continuo, situados en el maxilar inferior
y superior, y que solo están cubiertos de esmalte en la
parte frontal, y también, otros caracteres de los
esqueletos, como la separación de la tibia y la fíbula o
peroné, y la robustez del arco cigomático, que
observamos desde el Plioceno al Holoceno de Miramar.
Como se ha citado reiteradas
ocasiones, Sudamérica durante casi todo el Terciario,
quedo totalmente aislado del resto de los continentes,
en el cual, evolucionaron distintos grupos faunísticos
que no existían en otros continentes.
|
 |
Hay
evidencias, que, en varias oportunidades, entre
35 a 8 millones de años, antiguas balsas con
sedimentos, plantas y animales viajaron por el
océano atlántico desde África, cuando ambas
estaban separadas por apenas 900 kilómetros,
mientras que, otros grupos de animales (los
saltadores de islas) emigraban de la misma
forma, entre los archipiélagos aislados de lo
que hoy conforman el caribe, cuyo origen era
asiático, es lo que demuestran las pruebas
genéticas y paleontológicas. |
Una vez llegados a estas
tierras, se pudieron adaptar y diversificar fácilmente,
ocupando los nichos ecológicos de muchos notoungulados y
marsupiales, generando una importante competencia y
estrés ambiental.
Los primeros roedores de
América del Sur están representados por especies
descritas en depósitos del Eoceno/ Oligoceno temprano. A
finales del Oligoceno, todas las superfamilias y
familias de Caviomorpha estaban presentes en el registro
fósil.
En Miramar y gran parte del
Partido de General Alvarado, se han encontrado formas
muy diversas, e incluso, nuevas para la ciencia. Entre
los roedores coleccionados, tenemos el enorme Telicomys
giganteus, del tamaño de una vaca. Sus mandíbulas
presentan grandes incisivos, cuya cara anterior tienen
un ancho de 3 centímetros, acompañados por cuatro
molares por cada maxilar y rama mandibular. Suponemos
que su hábitat estaba limitado a lugares secos y con
follaje, aunque la evidencia fósil es muy escasa para
brindar datos más completos sobre su biología. Algunos
restos significativos fueron descubiertos en 1908 al
norte de Miramar por el propio Florentino Ameghino.
|
 |
Otro de los grandes
roedores Pliocenos encontrados en cierta
cantidad, lo conforma el Phugatherium novum,
emparentado con los actuales capibaras o
carpinchos, pero de mayor tamaño y patas largas
adaptadas para correr, y no de hábitos anfibios
como sus parientes actuales. Su cráneo refleja
un rostro alargado y estrecho. La longitud del
cráneo es la doble a la atribuida al carpincho,
superando fácilmente los 50 centímetros.
|
Su fémur y humero, guardan
la misma relación en su longitud con el género actual,
pero el cubito o ulna, radio, tibia, peroné y demás
huesos, son mucho más desarrollados y largos que estos,
por lo cual Phugaterium (=Protohydrochoerus),
morfológicamente concuerda con mamíferos corredores. En
el Pleistoceno, los fósiles de carpinchos son
extremadamente escasos, es tan así, que aún no sabemos
muy bien cuando aparecen los antecesores directos del
capibara viviente. Pero justamente en Miramar, se
encontraron huellas fosilizadas, atribuibles al
icnogenero Porcellusignum conculcator. De apariencia muy
similar a sus parientes sudamericanos modernos.
Actenomys latidens, fue un
mamífero roedor Caviomorpha muy común en la región
pampeana. Actenomys estaba emparentado con el actual
genero Ctenomys (tucos tucos) los cuales tienen una gran
diversidad en Sudamérica. Es probable que fuera de
hábitos subterráneos, ya que sus restos normalmente son
hallados en antiguas madrigueras, como así también,
Ctenomys (Paractenomys) chapadmalensis y Eucelophorus
cabrerai emparentados con Actenomys.
También había otros más
pequeños, como el Lagostomus (Lagostomopsis) antiquus,
es una especie de roedor de la familia Chinchillidae.
Actualmente sólo hay una especie viva, la vizcacha
común (Lagostomus maximus), que mide hasta 60
centímetros de largo y habita en Sudamérica, pero en el
Plioceno vivió Lagostomopsis, un representante de menor
tamaño.
|
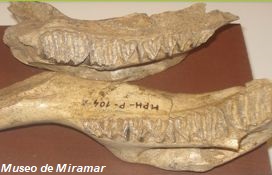 |
Su alimentación
estaría compuesta principalmente por tubérculos,
raíces, hierbas etc. Poseía unos cráneos anchos
y robustos, con nasales alargados. En la zona
norte de Miramar, en sedimentos del Plioceno,
entre 3,5 y 2,5 millones de años antes del
presente, se han recuperado numerosos restos de
Lagostomus (Lagostomopsis) incisus, asociado a
gran parte de la fauna extinta de fines de la
Era Terciaria. Además de restos óseos, se
observa largas madrigueras, en donde, en algunas
ocasiones se los han asociado a sus creadores.
|
Tambien, se han recuperado
integrantes de la familia Caviidae identificados (Dolichotis
sp, Dolicavia minuscula, Orthomyctera chapadmalensis,
Caviodon pozzii, Cardiomys sp., Microcavia
chapadmalensis, entre otros). Si bien muchos de ellos se
extinguieron, algunos representantes de cada familia
lograron sobrevivir desde el Plioceno al Holoceno
reciente. Entre las formas Pliocenas, encontramos a
Paleocavia impar, un género muy abundante durante el
Mioceno hasta el final del Plioceno. Como roedores
cavidos, pueden se indicadores de ambientes cálidos
templados. Tienen una cabeza grande y rectangular, así
como una cola muy pequeña o ausente. Poseen cuatro dedos
en las extremidades anteriores y tres en las
posteriores, de cuerpo alargado, y extremidades delgadas
y cortas, pero que sin embargo corre con gran facilidad
y rapidez. Vivía en lugares con vegetación abundante y
relativamente húmedos. Tenía unos 25 centímetros de
largo, y estaba relacionada con los conejillos de
Indias, cuises y el carpincho (el roedor viviente de
mayor tamaño).
Estos, son solo algunos
representantes fósiles que hemos encontrado en los
últimos 4 millones de años en nuestra zona, pero hay
muchísimos más de ellos para conocer. Miramar y zona, se
encuentra entre las localidades fosilíferas más ricas e
importantes del mundo, no solo por los gigantes del
pasado, sino también por los microvertebrados. Desde
estos sedimentos, se han organizado grandes colecciones
científicas sobre caviomorfos en los museos más
importantes del país, como así también Norteamérica y
Europa durante la primera mitad del siglo XX.
Bibliografía sugerida.
Aguirre, M. L. 1995. Cambios
ambientales en la región costera bonaerense durante el
cuaternario tardío, evidencias malacológicas. 4 jornadas
geológicas y geofísicas bonaerense. (Junín), actas 1:
35-45.
Candela, A., Cenizo, M.,
Tassara, D., Rasia, L., Robinet, C., Muñoz, N.,
Valenzuela, C. y Pardiñas, U. F. J. (2019).
A new echimyid genus (Rodentia,
Caviomorpha) in Central Argentina: uncovered diversity
of a Brazilian group of mammals in the Pleistocene.
Journal of Paleontology, 94 (1), 165-179.
Cenizo, M., Soibelzon, E. y Magnussen, M.
2015. Mammalian predator-prey relationships and
reoccupation of burrows in the Pliocene of the Pampean
Region (Argentina): new ichnological and taphonomic
evidence.
Historical Biology, 28 (8), 1026-1040.
Cione, A.L.; Tonni, E.P.
1995a. Bioestratigrafía y cronología del Cenozoico de la
región pampeana. In Evolución biológica y climática de
la región pampeana durante los últimos cinco millones de
años. Un ensayo de correlación con el Mediterráneo
occidental. Museo Nacional de Ciencias Naturales,
Consejo Superior de Investigaciones Científicas,
Monografías, p. 47-74. Madrid.
De los Reyes, L.M; Cenizo,
M.M.; Agnolin, F.; Lucero, S.; Bogan, S.; Lucero, R.;
Pardiñas, U.F.J.; Prevosti, F. & Scanferla, A. 2006a.
Aspectos paleofaunísticos y estratigráficos preliminares
de las sucesiones plio-pleistocénicas de la localidad
Centinela del Mar, provincia de Buenos Aires, Argentina.
90 Congreso Argentino de Paleontología y
Bioestratigrafía, 105.
Fidalgo, F. y Tonni, E.P.
1983. Geología y paleontología de los sedimentos
encausados del pleistoceno tardío y holoceno de Punta
Hermengo y arroyo Las Brusquitas (Partido de General
Alvarado y General Pueyrredon, Provincia de Buenos
Aires). Ameghiniana 20 (3-4): 281-296.
Genise, J.F. 1989. Las
cuevas con Actenomys (Rodentia, Octodontidae) de la
Formación Chapadmalal (Plioceno Superior) de Mar del
Plata y Miramar (Provincia de Buenos Aires). Ameghiniana,
26: 33-42.
Magnussen Saffer, M. 2005.
La Gran Extinción del Pleistoceno. Boletín de
divulgación Científico Técnico. Museo Municipal de
Ciencias Naturales Punta Hermengo de Miramar, Prov.
Buenos Aires, Argentina. Publicación 3: pp 3 – 10 (Sec.Pal).
Novas Fernando 2006. Buenos
Aires hace un millón de años. Editorial Siglo XXI,
Ciencia que Ladra. Serie Mayor.
Oliva, C. Boh, D. Magnussen, M
& Favier Dubois, C. 2018. Contribución al conocimiento
de Porcellusignum conculcator, Angulo y Casamiquela (Vertebratichnia,
Mammalipedia) del Cenozoico Superior (Mioceno Tardío –
Pleistoceno) de la Provincia de Buenos Aires
(Argentina). Jornadas Regionales VI Arqueológicas y VII
Paleontológicas Miramar. Libro de Resúmenes. Pag 33;34.
Olivares AI y Verzi DH. 2014.
Sistemática, filogenia y patrón evolutivo del roedor
hystricognath Eumysops (Echimyidae) del Plio -
Pleistoceno del sur de América del Sur. Biología
histórica, 2014
Olivares, A. I., Bastida, R.
O., Loza, C. M., Rodríguez, A. C., Desojo, J. B.,
Soibelzon, L. H. y
Quintana, C. A. 2008. Los
fósiles de Mar del Plata. Un viaje al pasado de nuestra
región. Buenos Aires, Argentina. Fundación de Historia
Natural “Félix de Azara”. 242 pp.
Vucetich
MG, Verzi DH. 1996. A new Eumysopinae (Rodentia,
Echimyidae) de la 'Formacion' Irene (Chapadmalalense
inferior?) Y la diversidad de la subfamilia. Actas IV
Jornadas geologicas geofisicas bonaerenses; Buenos
Aires, Argentina.
Tonni,
E.P.; Fidalgo, F. 1979. Consideraciones sobre los
cambios climáticos durante el Pleistoceno
tardío-Reciente en la Provincia de Buenos Aires.
Aspectos Ecológicos y
Zoogeográficos Relacionados. Ameghiniana, Vol. 15, Nos.
1-2, p. 235-253.
Los
fósiles de Perisodáctilos, que fueron hallados en
Miramar.
Por
Mariano Magnussen. Laboratorio Paleontológico del Museo
de Ciencias Naturales de Miramar. Fundación Azara. Laboratorio de
Anatomía Comparada y Evolución de los
Vertebrados.
marianomagnussen@yahoo.com.ar
. Ilustraciones Daniel Boh
Tomado de Magnussen,
Mariano. 2024. Los
fósiles de Perisodáctilos, en el Pleistoceno bonaerense.
Paleo Revista Argentina de Divulgación Paleontológica.
Boletín Paleontológico. Año XVII. 181: 57-60.
Son mamíferos ungulados que
se caracterizan por la posesión de extremidades con un
número impar de dedos terminados en una estructura
cornificada llamadas pezuñas, y con el dedo central, que
sirve de apoyo, mucho más desarrollado que los demás.
Los miembros de los
perisodáctilos se describen como mesaxónicos, es decir,
que su dedo central, en la mayoría de los casos el
tercero, el cual, se desarrolló mientras que los
restantes reducen su tamaño, hasta incluso desaparecer,
como los caballos.
 |
Se cree que los
perisodáctilos emergieron poco después de la
extinción masiva del Cretácico-Terciario en la
cual se extinguieron los dinosaurios y
otros organismos. Algunos tempranos hallazgos
datan del Paleoceno superior, entre hace 65 a 55
millones de años atrás, y que, para los inicios
del Eoceno, hace 54 millones de años, ya se
encontraban ocupando diversos continentes.
Inicialmente habitaron Asia y África, luego
fueron extendiéndose para ocupar Europa y
América del Norte. |
Durante todo la Era
Terciaria evolucionaron en estos continentes, pero
recién, en el Plioceno tardío, hace unos 2,8 millones de
años ingresaron al continente sudamericano, durante el
intercambio faunístico al unirse ambos subcontinentes.
A diferencia de otros
ungulados, como por ejemplo los rumiantes, conservan los
incisivos de la mandíbula superior y los utilizan para
arrancar hierba. Los dientes premolares se encuentran
molarizados, así junto a los molares forman una amplia
superficie para triturar el alimento.
Los équidos y tapíridos, son
las especies más emblemáticas entre los perisodáctilos,
sobrevivieron y prosperaron hasta el final
del Pleistoceno, hace solo 10 000 años, soportando la
presión de los cazadores humanos y de un nuevo hábitat
cambiante.
Los equinos, son mamíferos
placentarios del orden Perissodactyla, que contiene solo
un género viviente, Equus, y poseen en sus patas
un dedo modificado que soportan todo el peso, apoyado
sobre sus pezuñas. Estaban representados en el
Pleistoceno sudamericano, por Hippidion principale,
un primitivo caballo, el cual se extinguió hace unos 8
mil años.
 |
Era similar a las cebras de África, pero algo
más bajo y rechoncho, con extremidades cortas.
Su cráneo presenta unos huesos nasales algo
alargados y pronunciado que las formas
vivientes.
Por otro lado, es notable la conformación de las
extremidades, proporcionalmente cortas y anchas,
que le confieren al animal un aspecto macizo.
|
Su peso pudo ser de unos 400
kilos. Las características adaptativas de la parte
distal de las extremidades de Hippidion pueden
ser indicadores de distintos tipos de suelo y
vegetación. El hábitat de Hippidion debería
corresponder a un bosque o praderas húmedas y suelo
blando.
Los Équidos de América del
sur se han extinguido totalmente a principios del
Holoceno, y convivio con otra especie de caballo tardío,
el Amerhippus neogeus, un subgénero del género
Equus el cual agrupa a las cinco especies de dicho
género de la familia Equidae, que también se extinguió.
Recién, en el siglo XVI es
incorporado nuevamente el caballo a América, luego de la
colonización europea. En el museo se resguardan
distintos restos fósiles, y en exhibición se muestra un
cráneo muy completo con mandíbula de este extinto
animal.
Otro
perisodáctilo, el
Tapirus sp,
es un género de mamífero perisodáctilo extinto de la
familia de los tapíridos. El género Tapirus que
vivió en el Pleistoceno de América del Sur.
 |
Tapirus dupuy,
es una especie fue descrita originalmente por
Carlos Ameghino en 1916, empleando como base
material, fragmentarios colectados en sedimentos
del Pleistoceno de la ciudad de Miramar,
provincia de Buenos Aires, cuya especie
homenajea a José María Dupuy, prefecto local y
coleccionista en ciencias naturales, que
colaboro con la búsqueda y entrega de material
paleontológico y biológico al entonces Museo
Nacional de Historia Natural de Buenos Aires y
Museo de La Plata. |
Las extremidades anteriores
poseen cuatro dedos y las posteriores tres. Sin embargo,
la principal característica del Tapirus es su
alargado hocico en forma de pequeña probóscide, que usa
principalmente para arrancar las hojas, hierbas y raíces
que constituyen su alimento.
Los registros más antiguos
del género Tapirus en Sudamérica corresponden al
Pleistoceno temprano de Argentina, y probablemente de
Perú y Bolivia. Mientras tanto, en la actualidad, el
Tapir (Tapirus terrestres), que incluye a varias
subespecies que aun habita en el nordeste de Argentina,
casi todo Brasil, Bolivia, Paraguay, este de Ecuador y
Perú, las Guayanas, Venezuela y Colombia, es
descendiente de estas formas fósiles, que alguna vez,
habitaron la región pampeana durante la última edad de
hielo.
Bibliografia sugerida.
Alberdi Alonso M, Prado JL,
Favier-Dubois C. 2006. Nuevo registro de Hippidion
principale (Mammalia, Perissodactyla)
del Pleistoceno de Mar del Sur. Revista española de
paleontología, ISSN 0213-6937, Vol. 21, Nº. 2, 2006, págs. 105-114
Alberdi, M.T.; Prado, J.L.
1995b. Los Équidos de América del Sur. In Evolución
climática y biológica de la región pampeana durante los
últimos cinco millones de años. Un ensayo de correlación
con el Mediterráneo occidental (Alberdi, M.T.; Leone,
G.; Tonni, E.P.; editores). Museo Nacional de Ciencias
Naturales, Consejo Superior de Investigaciones
Científicas, Monografías, p. 295-308. Madrid.
Alberdi, M.T. & Prado, J.L.
2004. Caballos fósiles de América del Sur. Una historia
de tres millones de a-os. Incuapa, serie monográfica 3,
269 pp.
Ameghino,
F. 1888a. Rápidas diagnosis de algunos mamíferos fósiles
nuevos de la República Argentina, Buenos Aires, 17 pp.
Alberdi, G. Leone y E.P.
Tonni (eds)- Evolución biológica y climática de la
región pampeana durante los últimos cinco millones de
años. Un ensayo de correlación con el mediterráneo
occidental. Monografía del Museo Nac. Cs. Nat. Consejo
superior de investigaciones científicas. Madrid. pp.
77-104.
Bussing
WA, Stehli FG y Webb SD 1985. El gran intercambio
biótico estadounidense. Patrones de distribución de la
ictiofauna centroamericana, 453–473.
Cabrera, A., 1957. Catálogo
de los mamíferos de América del Sur. I. Rev. Mus. Cs.
Nat. “B. Rivadavia”, Zool. 4(1); 1-307. Bs.As.
Cione,
A.L. & Tonni, E.P. 2005. Bioestratigrafía basada en
mamíferos del Cenozoico superior de la región pampeana.
In: Geología y Recursos Minerales de la Provincia de
Buenos Aires (de Barrio, R.; Etcheverry, R.O.; Caballé,
M.F. & Llambías, E., eds.). XVI Congreso Geológico
Argentino, La Plata, Relatorio 11, 183-200.
Magnussen
Saffer, M. 2005. La Gran Extinción del Pleistoceno.
Boletín de divulgación Científico Técnico. Museo
Municipal de Ciencias Naturales Punta Hermengo de
Miramar, Prov. Buenos Aires, Argentina. Publicación 3:
pp 3 – 10 (Sec.Pal).
Novas Fernando 2006. Buenos
Aires hace un millón de años. Editorial Siglo XXI,
Ciencia que Ladra. Serie Mayor.
Olivares, A. I., Bastida, R. O., Loza, C.
M., Rodríguez, A. C., Desojo, J. B., Soibelzon, L. H. y
López, H. L. 2016.
Catalogue
of marine mammals of the mammalogical collection of the
Museo de La Plata, Argentina. Revista
del Museo de La Plata, 1: 57-82.
Quintana, C. A. 2008. Los
fósiles de Mar del Plata. Un viaje al pasado de nuestra
región. Buenos Aires, Argentina. Fundación de Historia
Natural “Félix de Azara”. 242 pp.
Sánchez B., Prado JL Alberdi
y MT .2006. Alimentación, ecología y extinción antiguas
de caballos del Pleistoceno de la Región Pampeana,
Argentina. Ameghiniana 43: 427–436
Tonni EP, Cione AL y Figini
AJ 1999. Predominio de climas áridos indicados por
mamíferos en las pampas de Argentina durante el
Pleistoceno tardío y el Holoceno. Paleogeografía,
Paleoclimatología, Paleoecología 147: 257–281.

|


























