|
Los
Grandes
Dinosaurios
Depredadores.
Fragmento del articulo publicado
originalmente: Magnussen Saffer, Mariano. (2009). Los
Grandes Dinosaurios Depredadores.
Paleo. A±o 7 numero 34: 11 - 13.
marianomagnussen@yahoo.com.ar
A
pesar de haberse encontrado menos de una docena de ejemplares
f¾siles, el
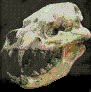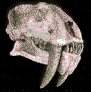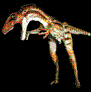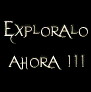
Tyrannosaurus
es uno de los dinosaurios mßs conocidos. En
1908, el paleont¾logo norteamericano Barnum Brown -que habĒa
realizado el primer descubrimiento de este animal, seis a±os antes
loŁgr¾ las mejores piezas rescatadas hasta ahora. Sin emŁbargo,
apenas dos o tres esqueletos completos se conŁservan en el mundo.
Con 5 metros de altura, este poderoso carnicero fue quizß el mayor
de los animales carnĒvoros que hayan pisado la tierra.
No optante, en Argentina se dio a conocer al Giganotosaurus
en 1996, y recientemente (2008) el
Tyrannotitan,
dos nuevos gigantes y depredadores carnĒvoros del Cretßcico que
superan en tama±o y fuerza al Tyranosaurus.
 |
El
Giganotosaurus
fue hallado por el Se±or Rubķn Carolini en las areniscas del
Miembro Candeleros de la formaci¾n Ri¾ Limay, que se exponen
en la zona de la represa de El Chocan, Provincia de Neuquķn,
Republica Argentina. Gran parte del esqueleto fue hallado,
incluyendo el crßneo, el cual, mide 1,8 metros de largo, un
numero importante de distintas vķrtebras, la pelvis, el
miembro posterior incompleto y algunas piezas de la cintura
espectacular. El tama±o de su crßneo nos demuestra que es el
carnĒvoro mas grande del mundo, lo cual es, por cierto, un
merito notable para el registro y estudio de los Dinosaurios
de nuestro paĒs y de Amķrica del sur. |
El fķmur de
Giganotosaurus mide 1,4 metros de largo, y como su posici¾n
era bĒpeda, o sea que caminaba erguido sobre sus patas traseras,
debi¾ medir aproximadamente unos 9 metros de alto hasta la cabeza y
unos 16 hasta la cola. Los estudios realizados en 1995 concluyeron
que se trata de una especie endķmica de Amķrica del sur, con
caracteres anat¾micos propios. Aunque la talla y su rol ecol¾gico
son comparables al del famoso Tyrannosaurus del
hemisferio norte, sus relaciones e historia filogenķticos son muy
distintas, considerßndolo como convergencia adaptativa o evoluci¾n
paralela, aunque podrĒan haber compartido ancestros comunes, que
luego siguieron ramas evolutivas aisladas geogrßficamente. De este
modo Tyrannosaurus fue producto de una evoluci¾n de
Laurasia, y Giganotosaurus de una evoluci¾n permanente
en Gondwana. Vivi¾ hace 95 millones de a±os.
 |
Por su lado
Tyrannotitan fue
presentado por el paleont¾logo
Fernando Novas. Lo llamaron asĒ porque era inmenso, llego a
medir unos 13 metros, carnĒvoro y camin¾ por el territorio
de la actual provincia de Chubut, Argentina. Eso fue 100
millones de a±os atrßs, durante el PerĒodo Cretßcico de la
Era Mesozoica. El Tyrannotitan
chubutensis
perteneci¾ a la familia de los cßrcarodontosaurios que s¾lo
se desarrollaron en el hemisferio sur. Junto con otras
especies, como los espinosaurios, los cßrcarodontosaurios se
habĒan extinguido unos 95 millones de a±os atrßs.
|
Sus signos distintivos eran sus dientes
afilados con peque±Ēsimas "monta±itas" y unas ciertas prominencias
en su lomo que estaban rodeadas de grasa. El clima era muy cßlido y
a·n los casquetes polares no existĒan cuando
Tyrannotitan chubutensis
asustaba por la Patagonia.
Los paleont¾logos conjeturan a partir de la forma y del ßngulo de
inserci¾n de sus huesos, por ejemplo el tipo de andar y la velocidad
que alcanzaban estos animales, muy veloces para su tama±o.
Lo que resulta evidente es que, a pesar de sus dimensiones, los
grandes depredadores tenĒan el cuerpo conformado en forma bastante
semejante al de los peque±os y ßgiles dinosaurios cazadores. Esto se
explica por la coincidencia de vida, pues ambos eran predadores. Y
esto permite comprender tambiķn la velocidad que alcanzaban en la
persecuci¾n de su caza. En cierto modo, entre las velocidades
logradas por los grandes carnĒvoros como
Tyrannosauru, Giganotosauru
y
Tyrannotitan,
y los peque±os ter¾podos existĒa una relaci¾n comparable a la que
hoy puede establecerse entre un veloz leopardo y un perro salvaje.
 |
Lo que sin duda llama la atenci¾n en las reconstrucciones de
estos ter¾podos es lo peque±o del miembro anterior,
ridĒculamente chico con relaci¾n a la tremenda cabeza y a
las fuertes patas del monstruo.
Ello es indicio de que en estos grandes carniceros los
miembros ya estaban extraordinariamente especializados para
la caza, lo que tambiķn puede observarse en los poderosos
predadores que vivieron en los perĒodos Jurßsico y
Cretßcico. |
El Allosaurus, por ejemplo, que con 12 metros de largo
fue el mßs grande de los dinosaurios carnĒvoros que habitaron la
Amķrica del Norte. Aunque no siempre este aniŁmal era carnĒvoro: se
piensa que, al menos en el caso de Apatosaurus
(gigantesco saur¾podo de 21 metros de largo), el Allosaurus
se alimentaba de su carro±a, ya que se encontraron marcas de dientes
en un esqueleto del saur¾podo. Y no parece probable que un
Apatosaurus vivo resultara fßcil presa para un
Allosaurus.
El gķnero Allosaurus, por otra parte, pudo ser
establecido ya en 1877, a partir de una serie de hallazgos
realizados en la Amķrica del Norte desde 1869, cuando apareci¾ un
primer resto f¾sil en las monta±as de Colorado. Estados Unidos.
Entre 1883 y 1884 pudo recuperarse un esqueleto casi completo, que
fue llamado Antrodemus en 1920. Mßs tarde se
estableci¾, sin embargo, que ese presunto nuevo gķnero no era sino
el mismo Allosaurus.
 |
El Carnotaunis sastrei, enconŁtrado en
los yacimientos de Cerro C¾ndor, en el Chubut, por el
paleont¾logo argentino Josķ Bonaparte. es uno de los escasos
dinosaurios teropodos de los cuales llegaron hasta nosotros
detalles de su rugosa piel. Su fuerte y corta cabeza,
ademßs, estaba protegiŁda por cuernos poco comunes,
seguramente muy ·tiles para defender los ojos en los duros
combates. Esta especie se pudo establecer a partir de dos
ejemplares incompletos, de los que surge una talla de
aproximadamente 4 a 5 metros de largo por 2 y medio de alto
hasta la cabeza, elevada por la posici¾n bĒpeda. El crßneo
debi¾ medir unos 60 centĒmetros de largo. Los fuertes
dientes, c¾nicos, estaban curvados hacia atrßs, seŁguramente
para arrancar la carne de la presa. |
Como en las aves corredoras actuales y en todos los dinosaurios
ter¾podos, las fuertes patas del Carnotaurus se
afirmaban sobre tres dedos.
Otro dinosaurio carnĒvoro hallado en la Argentina es el
Piatnitzkysaurus floresi, igualmente bĒpedo y de
brazo corto: tambiķn, un dinosaurio ya perfectamente especializado
para la caza. Es que, a pesar de su tama±o, los grandes dinosaurios
predadores demostraron estar entre los mßs feroces cazadores que
alguna vez galoparon el planeta. Los dinoŁsaurios carnĒvoros de
Amķrica del Sur, tuvieron un desaŁrrollo de formas independientes de
los otros grandes predadores que fueron encontrados en yacimientos
de Amķrica del Norte, de acuerdo a un fen¾meno que se llama
convergencia Evolutiva o Adaptativa. Este fen¾meno explica que
animales con conductas semejantes han evolucioŁnado de manera
diferente que sus "pares".
El genero Carcharocles desde el Eoceno al
Pleistoceno y su cronologĒa fosilifera.
Por Mariano Magnussen Saffer. Integrante de Grupo Paleo, Museo
Municipal Punta Hermengo de Miramar y Fundaci¾n Argentavis.
marianomagnussen@yahoo.com.ar
-
Magnussen Saffer,
Mariano (2014). El genero
Carcharocles desde el Eoceno al Pleistoceno y su cronologĒa fosilĒfera. Paleo,
Revista Argentina de PaleontologĒa. BoletĒn Paleontol¾gico. A±o XII. 101:
11-16.
Los
tiburones aparecen por primera vez en el registro f¾sil hace 455
millones de a±os. Sin embargo, no se vuelven comunes sino hasta
el Dev¾nico, hace unos 400 millones de a±os.
Estudiar la
evoluci¾n de estos animales marinos no es fßcil ya que al tener
esqueleto de cartĒlago, no se fosilizan en su totalidad.
Afortunadamente se han encontrado dentĒculos, dientes y
vķrtebras fosilizadas que nos permiten conocer los orĒgenes de
los tiburones.
El tibur¾n
mas antiguo es el
Antarctilmna
(que significa tibur¾n lamnoide de la antßrtica). Se ha
sugerido que este antiguo animal tiene unos 380 millones de a±os
y probablemente vivĒa en aguas dulces
Los
tiburones en sus formas
tempranas eran muy peque±os. Hay pruebas que sugieren que se parecĒa
mßs a los peces antes de
evolucionar a lo que son hoy. Muchos de ellos eran
similares a las anguilas. No tenĒan un cerebro tan grande como los
de hoy. Mßs de 2 / 3 del tama±o del cerebro de un
tibur¾n actual es para los sentidos, sobre todo su capacidad
para oler. Esta es una de las ßreas donde los
tiburones se han visto notablemente capaces de
evolucionar y
convertirse en una especie mßs inteligente.
Sus dientes
tambiķn eran muy diferentes a los de hoy dĒa. En lugar de tener gran
nitidez eran suaves. Sin embargo, estos primeros antepasados
definitivamente tenĒan las filas de dientes sustituibles como los
actuales. Esto podrĒa indicar que los antepasados de los tiburones
se alimentaban de plantas y que algo ocurri¾ e hizo imposible la
vida vegetal por un tiempo, con el fin de sobrevivir, empezaron a
consumir otros seres vivos,
evolucionaron.
Probablemente no
nadaban tan adentro en el ocķano como lo hacen hoy. Esto se
evidencia por la forma y el tama±o de sus aletas. Es posible que
hayan tenido que evolucionar
de esta manera para desplazarse largas distancias en
busca de alimentos.
 |
Un tibur¾n
pierde un diente, ķste cae y queda sobre el sedimento. El
diente puede desintegrarse (a) o enterrarse en el sedimento
(b); luego los procesos quĒmicos ocurren; aquĒ puede que el
f¾sil se disuelva (c), pero tambiķn puede ķste absorba los
minerales del sedimento o del agua en los espacios vacĒos
del diente y asĒ fosilizarse (d); ķste proceso dura en total
miles de a±os. El diente adquiere diferentes colores
dependiendo de los minerales que absorbi¾. Si la roca es
erosionada, despuķs de muchos a±os, es posible que
encontremos ķste diente f¾sil.
Otodus
obliquus
es una especie extinta de elasmobranquio lamniforme de la
familia Otodontidae, de gran tama±o, que vivi¾ durante el
Paleoceno y el Eoceno, hace entre 45 y 55 millones de a±os.
|
Otodus
obliquus
es, seg·n muchos paleont¾logos, el ancestro original del mßs grande
tibur¾n depredador que ha existido, Carcharodon megalodon.
Hay, sin embargo, otra escuela de pensamiento que sugiere que C.
megalodon fue en realidad descendiente de Carcharodon
orientalis (tambiķn conocido como Paleocarcharodon orientalis)
y este serĒa el predecesor de los modernos tiburones blancos
Carcharodon carcharias.
Aunque el
argumento de ambos puntos de vista es atractivo, la evidencia
muestra suficientes debilidades para justificar la precauci¾n en la
clasificaci¾n de C. megalodon en el mismo gķnero que el
tibur¾n blanco moderno (es decir, Carcharodon) y, por lo
tanto, ha sido reclasificado recientemente, para reflejar las
pruebas de montaje que Otodus obliquus probablemente fue el
antepasado de C. megalodon.
Nuestra
atenci¾n, sobre tiburones, se basa en el genero Carcharocles,
cuyos regiros fosilĒferos ocupan las capas sedimentarias desde el
Eoceno al Pleistoceno, con formas simples y corpulentas a terribles
monstruos de varias toneladas, representado por cuatro especies
fundamentales.
Carcharocles
auriculatus.
Jordan,
1923.
Carcharocles auriculatus es una especie extinta de tibur¾n incluida en el gķnero
Carcharocles, cercanamente relacionada con el mejor conocido
Carcharocles megalodon.
 |
Como
ocurre con muchos tiburones extintos, esta especie es
conocida a partir de dientes f¾siles y algunos centros de
vķrtebra. El esqueleto de los tiburones estß compuesto de
cartĒlago y no de hueso, material que raramente se fosiliza.
La longitud
de los dientes de C.auriculatus es relativamente
grande - de 25 a 114 milĒmetros. Sin embargo, es menor que
la de C. megalodon y Carcharocles angustidens.
Muchos de
los dientes de C. auriculatus provienen de Carolina
del Sur, Estados Unidos. Sin embargo, se conocen hallazgos
adicionales de dientes del Eoceno en la meseta Khouribga, en
Marruecos. Carcharocles auriculatus es el miembro mßs
primitivo del gķnero Carcharocles.
|
Carcharocles
angustidens. Agassiz, 1843
Carcharocles angustidens es una especie de tibur¾n megadentado extinto, que vivi¾
durante los perĒodos Oligoceno y Mioceno entre hace aproximadamente
35 a 22 millones de a±os. Se cree que este tibur¾n es un pariente
cercano del mßs conocido de los tiburones megadentados, C.
megalodon. Sin embargo, asĒ como en el caso de C. megalodon,
la clasificaci¾n de esta especie es materia de debate.
 |
Los restos
de C. angustidens por lo general estßn en muy mal
estado de preservaci¾n. A la fecha, el espķcimen mejor
preservado de esta especie ha sido excavado en Nueva
Zelanda, el cual abarca 165 dientes en asociaci¾n y cerca de
35 centros de vķrtebras asociados. Este espķcimen data de
hace cerca de 26 millones de a±os. Los dientes de C.
angustidens son notables por sus coronas triangulares y
sus peque±as c·spides laterales que estßn totalmente
aserradas. |
Los bordes
aserrados son muy afilados y muy pronunciados. C. angustidens
era una especie con una amplia distribuci¾n geogrßfica con f¾siles
hallados en Norteamķrica, Suramķrica, Europa, ┴frica, Nueva Zelanda,
Jap¾n, Australia, y Malta.
Igual que en
otros tiburones megadentados, los f¾siles de C. angustidens
indican que era considerablemente mayor que el actual gran tibur¾n
blanco, Carcharodon carcharias. Se estima que el espķcimen
bien preservado de Nueva Zelanda medĒa 9.3 metros de largo. Este
ejemplar tenĒa dientes que miden mßs de 9.87 centĒmetros en altura
diagonal, y centros de vķrtebras de cerca de 1.10 centĒmetros de
dißmetro. Sin embargo, existen reportes de f¾siles a·n mayores de
C. angustidens.
Carcharocles
chubutensis.
Es una especie
extinta de tibur¾n megadentado que vivi¾ durante los perĒodos
Oligoceno, Mioceno y Plioceno, entre hace 28 a 5 millones de a±os.
Este tibur¾n es considerado como unpariente cercano de otro tibur¾n
megadentado prehist¾rico, el famoso C. megalodon. Sin
embargo, como ocurre con C. megalodon, la clasificaci¾n de
esta especie estß en discusi¾n.
Esta especie es
conocida a partir de dientes f¾siles y algunos centros de vķrtebra.
El esqueleto de los tiburones se compone de cartĒlago y no de hueso,
y el cartĒlago raramente logra fosilizarse. Por lo tanto, los
f¾siles de C. chubutensis estßn generalmente mal preservados.
 |
Aunque
los dientes de C. chubutensis son morfol¾gicamente
similares a los de C. megalodon, estos son
relativamente delgados con una corona curvada, y con la
presencia de talones laterales apenas aserrados. Los f¾siles
de esta especie se han hallado en Norteamķrica, Sudamķrica,
Cuba, Puerto Rico, ┴frica, y Europa.
C.
chubutensis
era mayor que C. angustidens. Los dientes de C.
chubutensis se aproximaban a 130 milĒmetros de altura
perpendicular (longitud diagonal), la cual de acuerdo con el
mķtodo de estimaci¾n de tama±o propuesto por Gottfried at
al, in 1996, indica que el espķcimen tendrĒa una longitud de
12.2 metros. |
La investigaci¾n
paleontol¾gica sugiere que esta especie puede haber cambiado sus
preferencias de hßbitat a travķs del tiempo, o que pudo haber tenido
la suficiente flexiblilidad en su comportamiento para ocupar
distintos ambientes en distintas ķpocas.
C. chubutensis
era probablemente un superdepredador y cazarĒa generalmente peces,
tortugas marinas, cetßceos (por ejemplo, ballenas), y sirenios.
Carcharodon
megalodon.
Agassiz, 1843
es una especie
extinta de tibur¾n que vivi¾ aproximadamente entre 28 y 1,5 millones
de a±os atrßs, durante el Cenozoico (de finales del Oligoceno hasta
principios del Pleistoceno).
La asignaci¾n
taxon¾mica de C. megalodon se ha debatido por cerca de un
siglo, y a·n se encuentra en disputa con dos interpretaciones
principales: como Carcharodon megalodon (bajo la familia
Lamnidae) o bien como Carcharocles megalodon (bajo la familia
Otodontidae).
C. megalodon
es considerado como uno de los mayores y mßs poderosos depredadores
en la historia de los vertebrados. Los estudios sugieren que C.
megalodon lucĒa en vida como una versi¾n corpulenta del gran
tibur¾n blanco actual, Carcharodon carcharias, llegando a
alcanzar los 16 metros de longitud total. Los restos f¾siles indican
que este tibur¾n gigante tuvo una distribuci¾n cosmopolita, con
ßreas de crĒa en zonas costeras cßlidas. C. megalodon
probablemente tuvo una influencia muy importante en la estructura de
las comunidades marinas de su ķpoca.
C. megalodon
es representado en el registro f¾sil principalmente por dientes y
centros de vķrtebras.
 |
Como en
los demßs tiburones, el esqueleto de C. megalodon
estaba formado de cartĒlago mßs que de hueso; esto resulta
en una pobre preservaci¾n de los especĒmenes encontrados.
Sin embargo, los restos f¾siles de C. megalodon
indican que tenĒa los centros vertebrales densamente
calcificados.
Entre
las especies existentes, se considera que el gran tibur¾n
blanco es la mßs anßloga a C. megalodon. La carencia
de esqueletos f¾siles bien preservados de C. megalodon
ha forzado a los cientĒficos a basarse en la morfologĒa del
gran tibur¾n blanco para inferir su aspecto y estimar su
tama±o. |
Debido a los restos
fragmentarios, estimar el tama±o de C. megalodon se ha
convertido en un reto. Sin embargo, la comunidad cientĒfica reconoce
que C. megalodon superaba en tama±o al tibur¾n ballena (Rhincodon
typus).
En 1973, un
icti¾logo de Hawaii, John E. Randall, present¾ un mķtodo para
estimar la longitud total del gran tibur¾n blanco. El mķtodo
propuesto es representado en forma de un grßfico trazado, el cual
demuestra la relaci¾n entre la altura de la corona (la distancia
vertical de la hoja del diente desde la base de la parte de esmalte
del diente hasta su punta) del mayor de los dientes de la mandĒbula
superior del gran tibur¾n blanco y su longitud total.
Estos dientes
produjeron una longitud total de cerca de 13 metros. En 1991, dos
expertos en tiburones, Richard Ellis y John E. McCosker, se±alaron
un defecto en el mķtodo de Randall. De acuerdo a ellos, la altura de
la corona dental de los tiburones no necesariamente se incrementa en
proporci¾n a la longitud total del animal.
|
 |
Esta
observaci¾n llev¾ a nuevas propuestas para dar con mķtodos
mßs exactos para determinar el tama±o del gran tibur¾n
blanco y especies similares.
En
1996 tres cientĒficos ¢ Michael D. Gottfried, Leonard J. V.
Compagno y S. Curtis Bowman ¢ despuķs de haber realizado una
minuciosa investigaci¾n y escrutinio de 73 especĒmenes del
gran tibur¾n blanco, propusieron una relaci¾n linear
conservadora entre la mßxima altura del mayor de los dientes
superiores. El mayor diente de C. megalodon al
alcance de este equipo era un espķcimen de segundo diente
anterior superior, cuya mßxima altura era de 168 mm. Este
diente habĒa sido descubierto por Compagno en 1993, y
produjo una longitud total estimada en 15.9 metros.
|
Sin embargo,
rumores de dientes mayores de C. megalodon persistĒan en esa
ķpoca. La mßxima altura dental para este mķtodo se mide como una
lĒnea vertical desde la punta de la corona dental a la parte baja de
los l¾bulos de la raĒz, paralelo al largo eje del diente. Dicho de
otra forma, la mßxima altura del diente es la altura perpendicular.
En 2010, varios
investigadores de tiburones ¢ Catalina Pimiento, Dana J. Ehret,
Bruce J. MacFadden y Gordon Hubbell Ś estimaron la longitud total de
C. megalodon sobre la base del mķtodo de Shimada, produjo una
longitud total de 16.8 metros.
El tema de la
extinci¾n de C. megalodon estß a·n bajo investigaci¾n. Hace
cerca de 3.1 millones de a±os, ocurri¾ un importante evento
geol¾gico Ś el cierre del paso marĒtimo de Centroamķrica, el cual
caus¾ cambios significativos ambientales y en las faunas alrededor
del mundo. Estos cambios probablemente fueron responsables de la
extinci¾n de C. megalodon.
BibliografĒa
Sugerida;
Agassiz, L.
1833-1844. Recherches sur les Poissons Fossiles, V. 1-5.
Imprimerie de Petitpierre, NeuchŌtel, Suisse.
Bretton W. Kent (1994). Fossil Sharks of the Chesapeake Bay Region.
Egan Rees &
Boyer, Inc. 146 pßginas.
Bruner, John (1997).
½The "Megatooth" shark, Carcharodon megalodon╗ Mundo Marino Revista
Internacional de Vida Marina.
Consultado el 22-01-2012.
Cione A.L., E.P. Tonni y J.O. San Crist¾bal.
A
middle Pleistocene marine transgression in central-eastern
Argentina. Current Research in the Pleistocene 19: 16-18; 2002.
Lambert, Olivier; Giovanni Bianucci, Klaas Post, Christian de Muizon,
Rodolfo Salas-Gismondi, Mario Urbina and Jelle Reumer (1 de julio de
2010). ½The giant bite of a new raptorial sperm whale from the
Miocene epoch of Peru╗. Nature (Peru) 466 (7302): pp. 105¢108.
Gottfried M.D., Fordyce R.E (2001). ½An Associated Specimen of
Carcharodon angustidens (Chondrichthyes, Lamnidae) from the Late
Oligocene of New Zealand, with comments on Carcharodon
Interrelationships╗. Journal of Vertebrate Paleontology 21 (4):
pp. 730¢739.
Glikman, L.S., 1980.. Evolution of Cretaceous and Caenozoic Lamnoid
Sharks:3-247, pls.1-33. Moscow.
Jordan, D.S. & Hannibal, H., 1923. Fossil Sharks and Rays of the
Pacific Slope of North America. Bulletin of the Southern California
Academy of Sciences, 22:27-63, plates 1-9.
Marsili, Stefano; Giorgio Carnevale, Ermanno Danesea, Giovanni
Bianuccia, and Walter Landinia (March 2007).
½Early Miocene
vertebrates from Montagna della Maiella, Italy╗. Annales de
Palķontologie (Italy: Elsevier) 93 (1): pp. 27¢66.
Rudwick, M. J. S.
(1987) [1976]. El significado de los f¾siles. Episodios de la
Historia de la PaleontologĒa. Ciencias de la Naturaleza. Madrid:
Hermann Blume. pp. 347.
Wroe, S.; Huber, D.
R. ; Lowry, M. ; McHenry, C. ; Moreno, K. ; Clausen, P. ; Ferrara,
T. L. ; Cunningham, E. ; Dean, M. N. ; Summers, A. P. (2008).
½Three-dimensional computer analysis of white shark jaw mechanics:
how hard can a great white bite?╗.
Journal of
Zoology 276
(4): pp. 336¢342.
Un Recorrido por el Cenozoico de Espa±a.
Por el Bi¾logo Alfredo Castilla Wandosell. Fuente:
Asociaci¾n Cultural Paleontol¾gica Murciana 2005.
Despuķs de la gran debacle de finales del Mesozoico, las primeras
faunas de vertebrados terrestres
en el Paleoceno, estaban bßsicamente compuestas por mamĒferos de
peque±a talla. Entre ellos los multituberculados, algunos
marsupiales, ungulados primitivos y alg·n que otro primate. Tambiķn
aparece una amplia gama de mamĒferos placentarios primitivos de
hßbitos insectĒvoros. Todos ellos conformaban el n·cleo a partir del
cual, en el Eoceno, se originarĒa la mayor parte de ¾rdenes de
mamĒferos que hoy conocemos.
Cuando se inicia este perĒodo, Europa estaba constituida por un
conjunto de islas aisladas de los continentes mßs pr¾ximos, que
dieron lugar al desarrollo de faunas endķmicas en cada ßrea. En lo
que a nosotros nos concierne, Espa±a era un trĒo de islas: una isla
formada por Castilla, Portugal, Galicia y Asturias, una isla al sur
entre la isla anterior y el norte de ┴frica: el macizo Bķtico-Rife±o
y una tercera isla formada por Catalu±a, Baleares, C¾rcega y
Cerde±a. Entre estas islas estaba situado un peque±o mar interior:
la cuenca del Ebro.
|
 |
Durante la primera parte del Eoceno, algunas cuencas,
presentaban una flora de tipo tropical, parecidas a las que
actualmente se encuentran en la amazonia, con lianas y
abundante vegetaci¾n acußtica. Los ßrboles parecĒan poblados
por una variada gama de peque±os mamĒferos, como primates
del gķnero
Adapis,
parecidos a los actuales lemures y roedores primitivos
semejantes a las actuales ardillas y cuyos hßbitos debieron
de ser probablemente semejantes. |
En tierra se encontraban ya herbĒvoros de talla mediana e incluso
grande (Phenacodus),
grandes tapires (Lophiodon),
tambiķn primitivos depredadores (Proviverra),
del grupo de los creodontos, estos fueron de los primeros grupos de
mamĒferos placentarios que desarrollaron hßbitos carnĒvoros.
El clima cßlido y
h·medo que predomin¾ durante el Eoceno inferior-medio favoreci¾ la
eclosi¾n de una variada gama de formas vegetarianas de talla grande.
Entre ellas los representantes de los ķquidos, la familia que agrupa
a caballos y formas afines.
En el trßnsito del Eoceno medio- superior, hace unos 40 millones de
a±os, se produjo la transici¾n hacia un ambiente mßs ßrido, con una
clara estacionalidad y una vegetaci¾n mßs dura y abrasiva. Este
cambio climßtico estß asociado a un neto descenso de las
temperaturas. Estos condicionantes favorecieron la proliferaci¾n de
formas como
Pachynolophus
y Propaleotherium,
mejor adaptadas a la ocupaci¾n de espacios mßs abiertos. Los
paleotķridos, mejor dotados para las nuevas condiciones ambientales
del Eoceno superior, dieron lugar en Europa a una extraordinaria
profusi¾n de especies de todos los tama±os y tipos, destac¾ el
gķnero
Paleotherium.
Dentro del orden de los creodontos, destac¾ en el Eoceno superior el
gķnero
Hyaenodon.
Se trata de formas carnĒvoras, que durante el Eoceno superior y el
Oligoceno inferior ocuparon el nicho ecol¾gico que hoy ocupan los
cßnidos y las hienas. Tras el cambio faunĒstico que se oper¾ en el
trßnsito Eoceno-Oligoceno y durante el Oligoceno, los creodontos
fueron definitivamente desplazados por los carnĒvoros verdaderos.
Algunos creodontos, sin embargo, sobrevivieron a lo largo de todo el
Mioceno.
|
|
|
|
 |
A pesar de las fluctuaciones climßticas que ya se observan,
la selva oceßnica se mantuvo sin grandes variaciones hasta
que empieza el siguiente perĒodo, denominado Oligoceno. En
ese momento, asistimos sobre los continentes a uno de esos
cambios ambientales que han puntuado la historia de la
Tierra. AsĒ, la Antßrtida, se congela por primera vez desde
la era Primaria. |
|
|
|
Al acumularse importantes cantidades de agua s¾lida en el polo Sur,
se produjo un descenso general del nivel del mar (hasta 30 m), esto
provoc¾ uno de los fen¾menos de extinci¾n a nivel mundial mßs
importantes de la historia animal. A este fen¾meno se le denomina
TEE (Terminal Eocene Event), de manera que el archipiķlago que fue
Europa durante el Eoceno qued¾ de nuevo conectado al continente
asißtico. El ambiente se hizo mucho mßs seco y continental, con
fuertes contrastes estacionales. Las grandes extensiones de selva
h·meda dejaron paso a amplias praderas herbßceas. De esta forma, un
importante n·mero de nuevos inmigrantes invadieron el ßrea europea,
determinando la extinci¾n de muchas de las formas que habĒan
predominado durante el Eoceno. A esto se le denomin¾ la "ruptura
faunĒstica de Stehlin".
Las primeras vĒctimas fueron los mamĒferos adaptados al bosque, como
los primates del tipo (Adapis).
Por lo que hace a los paleotķridos del Eoceno, la mayor parte se
extinguieron y tan s¾lo algunas especies de talla peque±a o mediana
sobrevivieron todavĒa durante la parte inferior del Oligoceno. En su
lugar, llegan desde Asia, los primeros rinocerontes. Entre los
artiodßctilos tambiķn tienen lugar una importante renovaci¾n.
A principios del Mioceno, hace unos 20 millones de a±os, la placa
africana, compuesta por ┴frica y Arabia se encontraba completamente
aislada de Europa y Asia por un ocķano transversal denominado Tethys.
En consecuencia, las faunas terrestres de estas masas continentales
estaban constituidas por grupos aut¾ctonos que habĒan evolucionado
independientemente durante millones de a±os. Pero durante el Mioceno
inferior, la placa tect¾nica africana colision¾ con Asia a nivel del
Pr¾ximo Oriente, y ello permiti¾ un importante intercambio
faunĒstico en ambos sentidos.
Algunos yacimientos de la penĒnsula muestran la tĒpica asociaci¾n de
inmigrantes africanos junto a formas persistentes del Oligoceno,
propia de estas faunas del Mioceno inferior. AsĒ, entre los
elementos de origen africano se encuentran mastodontes, pangolines
...Tambiķn encontramos a los denominados "perros oso", ciervos,
osos, suidos, b¾vidos, rinocerontes acußticos...
|
|
|
|
 |
El trßnsito del Mioceno inferior al Mioceno medio, hace unos
15 millones de a±os, marc¾ un profundo cambio ambiental en
la penĒnsula Ibķrica. De un biotopo h·medo y boscoso, con
una alta diversidad de especies, se pas¾ a un biotopo mucho
mßs seco y abierto. Las floras de esta ķpoca, muestran una
alta proporci¾n de plantas arbustivas de hoja peque±a (hasta
el 70 % principalmente mimosßceas). Ello indica que la
estaci¾n seca era mucho mßs prolongada que en ķpocas
anteriores. |
|
|
|
Los bosques, dominados por mimosas y acacias, se habrĒan reducido a
peque±os n·cleos cerca de los cursos de agua. La fauna de mamĒferos
estaba dominada por grandes herbĒvoros adaptados a una dieta basada
en arbustos. Al mismo tiempo, se produjo una drßstica reducci¾n de
la diversidad faunĒstica, especialmente en lo que respecta a las
especies asociadas al bosque. El final del Mioceno medio y el inicio
del Mioceno superior, hace unos 12 millones de a±os, marca en la
penĒnsula Ibķrica el retorno al biotopo h·medo y boscoso que
caracterizara el Mioceno inferior. Prueba de ello son los numerosos
restos de castores que aparecen en algunos yacimientos de esta
ķpoca. Por entonces las copas de los ßrboles eran surcadas por
diversas especies de ardillas voladoras como las que actualmente
pueblan los bosques del sudeste asißtico. Vamos tambiķn a
encontrarnos una gran profusi¾n de ciervos de diverso tipo.
SeguĒamos
encontrando mastodontes de gran tama±o, los suidos (parientes de los
actuales jabalĒes) volvĒan a ser elementos dominantes en los bosques
de este perĒodo. Los rinocerontes acußticos y de bosque vuelven a
ser dominantes, sustituyendo a las formas corredoras del tipo
Hispanotherium.
|
 |
Hace unos 10 millones de a±os se produjo un nuevo descenso
del nivel del agua a nivel global. Los ocķanos descendieron
cerca de 100 metros, de manera que nuevos puentes
intercontinentales emergieron, permitiendo la migraci¾n y el
intercambio entre las faunas terrestres. Este fue el caso de
Hipparion,
peque±o caballo de tres dedos, que sustituy¾ a los caballos
venidos ya desde Amķrica (Anchitherium).
|
|
|
|
A partir de este momento, los restos de
Hipparion
se hacen tan frecuentes en los yacimientos europeos del Mioceno
superior y del Plioceno, que este gķnero se ha convertido en un
f¾sil caracterĒstico de estas edades. En su migraci¾n, las primeras
oleadas de hippariones arrastraron consigo a otros grandes mamĒferos
de origen asißtico que acompa±aron a estos ķquidos en su viaje hacia
Europa, tal es el caso de los denominados "tigres diente de sable".
Junto a ellos, se encontraban tambiķn los primeros jirßfidos que
entraron en Europa. Hace unos 6 millones de a±os, se inicia la
desecaci¾n del mar Mediterrßneo, este hecho, abri¾ nuevos puentes de
comunicaci¾n entre las faunas terrestres de Europa, ┴frica y Asia.
El mar Mediterrßneo qued¾ reducido a unos pocos lagos salinos que
dieron lugar a las famosas "evaporitas". Para los aventureros,
comentar que este perĒodo de tiempo puede ser estudiado en Lorca,
donde podemos hallar una rica fauna piscĒcola propia de esta ķpoca.
En realidad, todo comenz¾ hace unos 7 millones de a±os en la zona
del estrecho de Gibraltar. En esta ķpoca, el estrecho como tal no
existĒa todavĒa. Un conjunto de arcos de islas, bordeadas por
arrecifes de coral, se extendĒan entre el norte de ┴frica y la
penĒnsula Ibķrica, en lo que hoy es el valle del Guadalquivir y la
costa de Alicante y Murcia. La progresiva deriva de placa africana
provocaba la aparici¾n de relieves cada vez mßs abruptos en este
zona. Pero el hecho determinante fue otro. El continente antßrtico
se vio sometido a una primera fase de glaciaci¾n. Este hecho tuvo
como efecto inmediato un descenso generalizado del nivel de los
ocķanos. Y como consecuencia, el ocķano Atlßntico dej¾ de estar
comunicado con el mar Mediterrßneo. Cuando hace 6 millones de a±os
se cort¾ la comunicaci¾n con este ocķano, un rßpido proceso de
desecaci¾n lo dej¾ convertido en una estepa cßlida, moteada en sus
partes mßs profundas por grandes lagos salobres parecidos al actual
mar Muerto. Desde el fondo del mar desecado, relieves como las
Baleares o C¾rcega y Cerde±a aparecĒan como extraordinarias
cordilleras en medio de un desierto salino.
|
|
|
|
 |
Entre los nuevos inmigrantes africanos del Messiniense
tambiķn se encuentran los grandes mamĒferos como los
hipop¾tamos, que hasta entonces eran elementos desconocidos
en el registro euroasißtico. Pero sin duda, uno de los
elementos mßs sorprendentes que entran durante el
Messiniense en la penĒnsula Ibķrica es
Paracamelus,
un tĒpico representante de la familia de los camellos. Este
gķnero ha sido encontrado en diversos yacimientos del
Mioceno terminal del Levante ibķrico, como Venta del Moro
(cerca de Valencia) y Librilla en Murcia.
|
El inicio del Plioceno comporta un importante cambio ambiental,
marcado por lo que se podrĒa calificar de nuevo ¾ptimo climßtico.
Las temperaturas medias anuales subieron de nuevo, situßndose entre
3 y 10 ║C por encima de las actuales. Por ·ltima vez en su historia,
la Antßrtida se vio de nuevo libre de hielo, desarrollando una
vegetaci¾n de tipo templado-subtropical. Como consecuencia, el nivel
general de los ocķanos subi¾ cerca de 60 metros, cubriendo muchas
ßreas que hasta entonces habĒan estado por encima del nivel de las
aguas. Los antiguos golfos y ca±ones desarrollados por la regresi¾n
del mar en el Messiniense, se vieron inundados ahora, formando
fiordos y estuarios.
Los yacimientos encontrados de este perĒodo muestran una composici¾n
faunĒstica que refleja las condiciones de ¾ptimo climßtico referida.
AsĒ, entre la fauna de grandes mamĒferos reaparecen los tapires. Las
tortugas gigantes del gķnero
Cheirogaster,
de mßs de dos metros, tambiķn hacen su aparici¾n frecuentando las
marismas. Reaparecen los primates, representados por macacos. Entre
los carnĒvoros se opera una importante renovaci¾n. AsĒ, aunque
persisten los grandes fķlidos, los elementos dominantes son ahora
los cßnidos. Los osos, castores y las ardillas voladoras tambiķn
estßn representados.
|
 |
Entre los elementos del Mioceno superior
Hippari¾n
contin·a siendo el herbĒvoro mßs abundante, aunque su
diversidad ha disminuido. Tambiķn son frecuentes los
mastodontes del gķnero
Anancus,
que a partir de este momento constituirßn los proboscĒdeos
dominantes de las faunas del Plioceno. Entre los rumiantes,
persisten en la parte mßs baja del Plioceno los ·ltimos
jirßfidos. Por el contrario, las formas de origen africano
que entraron durante el Messiniense en la penĒnsula Ibķrica,
como los hipop¾tamos o los camellos del gķnero
Paracamelus,
no sobreviven al trßnsito Mioceno-Plioceno. |
Hace unos 2'5 millones de a±os se produjo un nuevo enfriamiento a
nivel global, que determin¾ la aparici¾n de un primer casquete de
hielo en el ┴rtico y no s¾lo en la Antßrtida, como hasta entonces
habĒa ocurrido. Este cambio climßtico fue mucho mßs importante que
los anteriores, ya que con ķl se inici¾ el ciclo de perĒodos
alternantes glaciar-interglaciar que ha caracterizado a nuestro
planeta desde entonces. En este momento se detecta en Eurasia una
importante regresi¾n de las masas boscosas y la expansi¾n de las
praderas de gramĒneas. En ┴frica, el desierto del Sahara se
consolida como una barrera permanente, en tanto que el bosque
cerrado de ßfrica oriental entra en regresi¾n y se transforma en
amplias sabanas.
Aunque esta primera crisis climßtica no fue tan fuerte como las que
posteriormente se desarrollaron a lo largo del Cuaternario, sus
efectos se hicieron sentir sobre las faunas terrestres. AsĒ, en el
trßnsito al Plioceno superior se detecta la desaparici¾n de aquellos
elementos de tipo subtropical que, como los jirßfidos o los tapires,
estaban todavĒa presentes en el Plioceno inferior. A su vez, nuevos
inmigrantes de origen oriental desplazaron a muchos de los elementos
que habĒan persistido a lo largo del Plioceno. Los primeros
elefantes verdaderos, precursores de los elefantes actuales y del
Mamut del Pleistoceno, substituyeron, tras un corto intervalo de
coexistencia, a Anancus y a los ·ltimos mastodontes. Entre los
ķquidos, los ·ltimos hippariones desaparecieron definitivamente del
registro europeo tras la llegada de los primeros caballos
verdaderos.
|
|
|
|
 |
La fauna de grandes herbĒvoros se vio asimismo enriquecida
por la entrada de otros nuevos elementos; por ejemplo,
aparece un nuevo tipo de gacela, aparecen los primeros
b¾vidos de gran tama±o y porte pesado, los ciervos
experimentaron una importante diversificaci¾n. A este
respecto, decir que durante todo el Mioceno superior y hasta
el Plioceno inferior, los ciervos habĒan estado relegados a
unas pocas especies de talla peque±a y astas simples. Con la
crisis climßtica del Plioceno medio, el panorama cambia por
completo. |
|
|
|
Nuevos ciervos de mayor talla y astas complicadas entran en escena.
La dispersi¾n de estos grandes herbĒvoros estß sin duda relacionada
con la expansi¾n de las praderas herbßceas hace 2,5 millones de
a±os. El cuadro de los rinocerontes tambiķn vari¾ substancialmente a
partir del Plioceno superior, ya que en el Mioceno superior suponĒan
un grupo relativamente marginal, es en este momento cuando empiezan
a ser abundantes los representantes del gķnero
Stephanorhinus.
Sus representantes mßs primitivos,
Stephanorhinus
etruscus,
eran formas muy parecidas a los actuales rinocerontes africanos. En
Caravaca y Cartagena se han encontrado restos de estos animales.
Este cambio climßtico afect¾ tambiķn a las faunas de carnĒvoros. En
particular, la representaci¾n de este grupo a partir del Plioceno
superior se caracterizarß por la presencia de muchos de los gķneros
que hoy encontramos en el Viejo Mundo. AsĒ, junto a los primeros
linces, encontramos tambiķn elementos mßs ex¾ticos como los
guepardos y formas precursoras de la hiena rayada de la India.
Asociaciones de este tipo son caracterĒsticas de diversos
yacimientos del Plioceno superior ibķrico. Ello comport¾ asimismo la
desaparici¾n de otros carnĒvoros de origen miocķnico que habĒan
sobrevivido hasta el Plioceno inferior. Curiosamente, esta
renovaci¾n no afect¾ a los felinos "dientes de sable" presentes en
Europa desde el Mioceno superior. Por el contrario, a partir del
Plioceno superior este grupo aumenta incluso su diversidad con
respecto a etapas anteriores.
Mķtodo CientĒfico: Conocimientos bßsicos para la
realizaci¾n de una publicaci¾n cientĒfica.
Fragmento de:
Por Mariano
Magnussen Saffer. Director del Grupo Paleo y Presidente de la
Asociaci¾n Amigos del Museo Punta Hermengo.
marianomagnussen@yahoo.com.ar
El mķtodo
cientĒfico es un proceso destinado a explicar fen¾menos, establecer
relaciones entre los hechos y enunciar leyes que expliquen los
fen¾menos fĒsicos y naturales del mundo y permitan obtener, con
estos conocimientos, aplicaciones ·tiles al hombre. Los cientĒficos
emplean el mķtodo cientĒfico como una forma planificada de trabajar.
Sus logros son acumulativos y han llevado a la Humanidad al momento
cultural actual. Toda investigaci¾n cientĒfica se somete siempre a
una "prueba de la verdad" que consiste en que sus descubrimientos
pueden ser comprobados, mediante experimentaci¾n, por cualquier
persona y en cualquier lugar, y en que sus hip¾tesis son revisadas y
cambiadas si no se cumplen.
El mķtodo
cientĒfico (del griego: -meta = hacia, a lo largo- -odos = camino-;
y del latĒn scientia = conocimiento; camino hacia el conocimiento)
presenta diversas definiciones debido a la complejidad de una
exactitud en su conceptualizaci¾n: "Conjunto de pasos fijados de
antemano por una disciplina con el fin de alcanzar conocimientos
vßlidos mediante instrumentos confiables", "secuencia estßndar para
formular y responder a una pregunta", "pauta que permite a los
investigadores ir desde el punto A hasta el punto Z con la confianza
de obtener un conocimiento vßlido". AsĒ el mķtodo es un conjunto de
pasos que trata de protegernos de la subjetividad en el
conocimiento.
La fuente de esta
necesidad de conocer y de saber surge de su curiosidad, elemento
fundamental en la personalidad del investigador que lo lleva a
cuestionar, a indagar y por ende a adquirir los conocimientos que le
permitan evolucionar y trascender. De hecho, el conocimiento que le
ha sido legado a la humanidad y por el cual se ha logrado el
desarrollo que se vive en la actualidad, tiene como base las
investigaciones y descubrimientos que se encuentran plasmados en la
historia de la humanidad
Por proceso o
"mķtodo cientĒfico" se entiende aquellas prßcticas utilizadas y
ratificadas por la comunidad cientĒfica como vßlidas a la hora de
proceder con el fin de exponer y confirmar sus teorĒas. Las teorĒas
cientĒficas, destinadas a explicar de alguna manera los fen¾menos
que observamos, pueden apoyarse o no en experimentos que certifiquen
su validez. Sin embargo, hay que dejar claro que el mero uso de
metodologĒas experimentales, no es necesariamente sin¾nimo del uso
del mķtodo cientĒfico, o su realizaci¾n al 100%.
Historia.
Frente a los
lĒmites del azar o la casualidad que en pocas ocasiones nos dan
conocimiento, -ya sea conocimiento cientĒfico, del bien o, como
indica Arist¾teles en la ╔tica a Nic¾maco, del bien mßximo
que es la felicidad-, Plat¾n y el mismo Arist¾teles advertĒan de la
necesidad de seguir un mķtodo con un conjunto de reglas o axiomas
que debĒan conducir al fin propuesto de antemano. S¾crates, Plat¾n y
Arist¾teles, entre otros grandes fil¾fosos griegos propusieron los
primeros mķtodos de razonamiento filos¾fico, matemßtico, l¾gico y
tķcnico.
|
 |
Durante la
ķpoca medieval serßn los fil¾sofos, fĒsicos, matemßticos,
astr¾nomos y mķdicos del mundo islßmico quienes hagan suya,
desarrollen y difundan la herencia de la filosofĒa griega.
Se debe reconocer, entre otros, a figuras como Alhazen, Al-Biruni
y Avicena. Tambiķn debemos mencionarse a quines
contribuyeron a la difusi¾n de dichos conocimiento por
Europa; figuras como Roberto Grosseteste y Roger Bacon junto
con la imprescindible labor de Escuela de Traductores de
Toledo. Pero no serß hasta la edad moderna cuando se
consolide una nueva FilosofĒa Natural. Descartes (1596-1650)
en su obra el Discurso del mķtodo define por primera vez
unas reglas del mķtodo para dirigir bien la raz¾n y buscar
la verdad en las ciencias. |
A·n con
diferencias notables fueron muchos los que defendieron la necesidad
de un mķtodo que permitiera la investigaci¾n de la verdad.
Desde un punto
de vista empĒrico o cientĒfico tal y como ahora lo entendemos se
debe mencionar a precursores del mķtodo cientĒfico como Leonardo da
Vinci (1452-1519), Copķrnico (1473-1543), Kepler (1571-1630) y
Galileo (1564-1642) quienes aplicaban, con mayor o menor conciencia
una reglas met¾dicas y sistemßticas para alcanzar la verdad. Galileo
Galilei contribuy¾ a reforzar la idea de separar el conocimiento
cientĒfico de la autoridad, la tradici¾n y la fe.
Importancia.
El mķtodo se
encuentra entre la teorĒa y la realidad; gracias a ķl la
investigaci¾n cientĒfica puede realizarse y la ciencia continuar su
evoluci¾n y desarrollo. Por eso se dice que el mķtodo es importante,
es el camino que nos lleva hacia la verdad de las cosa. Generalmente
el hombre com·n considera ciencia al c·mulo de conocimientos, sin
embargo, hay que recordar que al conocimiento se le llama
ōcientĒficoö s¾lo porque ha sido conocido por el mķtodo cientĒfico,
o por lo menos ha sido adquirido por medio del mķtodo cientĒfico o a
sido puesto a prueba por ķl.
ōEl
conocimiento cientĒfico sirve, de hecho es el ·nico medio
disponible, para conocer el mundo real, es decir, es una de las
formas que tiene el hombre para otorgarle un significado con sentido
a la realidad.
ōEn el proceso
de producci¾n del conocimiento basado en la ciencia, los
planteamientos y teorĒas deben ser formulados de una manera rigurosa
y explĒcita estableciendo sus bases con criterios objetivosö. ōEl
proceso de producci¾n del conocimiento basado en la ciencia, no se
puede reducir a una colecci¾n de hechos, es por ello necesario
seleccionar los hechos y organizarlos de una manera sistemßticaö.
Es decir, que
debemos ser rigurosos en la formulaci¾n de los enunciados y muy
precisos en las metodologĒas que aplicaremos en el proceso de
nuestra investigaci¾n, o dicho de otra manera, que debemos ser
sistemßticos. Podemos entonces afirmar que hay conocimiento
cientĒfico, cuando a travķs del mķtodo cientĒfico, se ha logrado
acumular nuevos conocimientos, nuevas experiencias. Por lo tanto, la
ciencia avanza en la medida en que se logren plantear y resolver los
problemas, es decir, el progreso del conocimiento se da en la medida
en que se descubren y se resuelvan nuevas dificultades.
Puntos fundamentales para la
utilizaci¾n del Mķtodo CientĒfico en un trabajo de Investigaci¾n.
TĒtulo; El tĒtulo es la primera
parte de un artĒculo. Debe ser corto, especĒfico e informativo. En
lo posible debe brindarle al lector la debida informaci¾n de su
contenido: Su naturaleza del estudio, organismo experimental, lugar
y perĒodo de tiempo del estudio y enfoque metodol¾gico.
Autores; En general el orden de
los autores del trabajo esta en funci¾n de la participaci¾n (por ej.
dedicaci¾n, etc . ), Deben mencionar instituci¾n donde desarrollan
sus actividades o relaci¾n con el estudio, direcci¾n postal o correo
electr¾nico para que otros investigadores puedan contactarse con los
autores.
Resumen, summary o abstract; Es
por lo general lo que se lee a continuaci¾n del tĒtulo. Debe decirle
al lector de quķ trata el artĒculo. Contiene un resumen de los
contenidos y conclusiones. Remarca cualquier informaci¾n nueva e
indica la relevancia de la misma. Debe ser lo suficientemente
explicativo como para que la naturaleza, objetivos y logros del
trabaje puedan ser entendidos. Algunos consejos pueden ser ·tiles a
la hora de elaborar un resumen, como son: usar oraciones completas,
evitar el uso de jerga, no abreviar las palabras, omitir en la
medida las referencias y describir en forma resumida la metodologĒa
utilizada. Recordar que un buen resumen puede motivar a un lector a
solicitar el trabajo a su autor. Muchas veces buenos trabajos quedan
sin leerse por que el resumen estß mal elaborado y no da la
suficiente informaci¾n.
Introducci¾n; La introducci¾n
esta compuesta por dos partes: una son los antecedentes y en la otra
el objetivo del trabajo. En los antecedentes se debe priorizar toda
aquella informaci¾n de otros autores que luego van a ser discutida
en la Discusi¾n, en general confrontßndola con nuestros resultados.
El o los objetivos del trabajo deben ser bien explicitados y en
general deben ser ubicados al final de la introducci¾n.
Materiales y Mķtodos; Esta es
una de las secciones mßs importantes de un trabajo cientĒfico. Uno
de postulados bßsicos de la actividad cientĒfica es que los
resultados obtenidos deben repetibles. Por lo tanto esta secci¾n
debe ser lo suficientemente clara y detallada para que otro
investigador en cualquier lugar del mundo pueda repetir el trabajo.
Los puntos que conviene tener en cuenta al redactar materiales y
mķtodos son: los aparatos, instrumentos de origen comercial s¾lo
deben ser nombrados, los aparatos o instrumentos hechos a medida
deben ser descriptos en detalle y si alguno de los mķtodos usados ha
sido descripto previamente s¾lo se lo puede citar.
Resultados; Se utilizan
aquellos resultados que son pertinentes al trabajo. En general se
recomienda presentar los resultados en un formato narrativo que
utilice tablas y figuras que corroboren las afirmaciones realizadas.
Las tablas y figuras deben presentar tĒtulos claros, informativos y
completos. Siempre se debe recordar que los resultados son del
trabajo realizado, por lo tanto en esta secci¾n se debe evitar hacer
referencia a otros autores.
|

|
Discusi¾n; Recordar que en la
discusi¾n se deben discutir los resultados propios y
aquellos obtenidos por otros autores, en particular aquellos
ya mencionados en los antecedentes de la introducci¾n. Una
buena discusi¾n jerarquiza un trabajo, por lo que es_ aquĒ
donde el investigador conjetura y confronta sus propios
resultados con otros colegas y ademßs puede plantear
hip¾tesis creativas sobre problemas a·n no resueltos y
orientar la futura investigaci¾n en el tema.
Conclusiones; Son las
respuestas a los objetivos del trabajo, por lo tanto se debe
utilizar expresiones claras, concisas y contundentes.
|
Agradecimientos; Es una secci¾n
que se acostumbra a incluir en los trabajos de investigaci¾n. ┐A
quiķn se agradece? En general a persona que ayudaron en el trabajo,
en la lectura crĒtica del trabajo, como asĒ tambiķn a instituciones
que Financiaron total o parcialmente el mismo.
BibliografĒa Citada; En general
aparecen en orden alfabķtico por el apellido del autor. El estilo en
las citas difiere de una publicaci¾n a otra, por lo cual el autor
debe informarse de las normas de publicaci¾n de la revista donde
desea publicar su artĒculo. Por lo general para una publicaci¾n
peri¾dica el orden de presentaci¾n es: autor/es, - a±o o fecha de la
publicaci¾n - tĒtulo del artĒculo - nombre de la publicaci¾n -
n·mero del volumen v n·meros de pßginas.
Mķtodo 3 R; Evaluaci¾n de un
trabajo cientĒfico.
Habiendo estructurado el
trabajo en funci¾n de las pautas desarrolladas ahora corresponde
determinar si existe alguna metodologĒa rßpida, clara y eficiente
para poder juzgarlo y ademßs tener una herramienta que nos permita
analizar si hemos ubicado cada Ētem en el lugar correspondiente.
Apelamos a una estrategia que hemos denominado las 3 R, y que son:
la Relaci¾n, la Respuesta y el Respaldo.
La Relaci¾n entre TĒtulo del
trabajo y el/los Objetivo/s que nos hemos
propuesto. Por ejemplo: TĒtulo: "
Distribuci¾n geogrßfica y ecol¾gica de Felis concolor
en la regi¾n pampeana". Objetivo: " El objetivo de este
trabajo es cuantificar la presencia del genero y especie en cuesti¾n
en distintos ambientes, tanto naturales como artificiales y su
relaci¾n con el medioö.
Respuesta de los Objetivos en
la/s Conclusi¾n/es. Es recomendable que en el
trabajo figuren explĒcitamente las Conclusiones a las
que se llegaron, esto facilita la lectura y evaluaci¾n del mismo.
En muchas publicaciones figura Discusi¾n y Conclusi¾n juntos, pero
es preferible que estķn separados porque la Discusi¾n,
desde un punto de vista metodol¾gico y epistemol¾gico
crucial y quizßs lo mßs importante de un trabajo de investigaci¾n y
junfarla con las conclusiones no es
recomendable.
Respaldo de la/s Conclusi¾n/es
en los Resultados. En las ciencias naturales
(fĒsica, quĒmica y biologĒa y afines) es fundamental el trabajo
empĒrico y experimental, por lo tanto esta R es crucial a la hora de
evaluar Metodol¾gica y epistemol¾gicamente un manuscrito. La
credibilidad de la ciencia empĒrica estß en el apoyo brindado por la
realidad y aunque esto tenga claras implicancias empiristas (como
corriente filos¾fica) no nos queda mßs remedio que a los hechos para
justificar nuestros resultados.
BibliografĒa
Sugerida:
Bunge, M. 1977. " ┐ Quķ es la
ciencia ? ". pp. 9-36. " ┐ Cual es el mķtodo de la ciencia ? ". pp.
39-68. En: La ciencia: su mķtodo y su filosofĒa. Ediciones SIGLO
XX
F═A.
Bunge, M. 1981. " ┐ Quķ es y a
quķ puede aplicarse el Mķtodo CientĒfico ? ". En: EpistemologĒa, pp.
28-45. Editorial ARIEL.
Chalmers, A. 1990. " El
inductivismo: la ciencia como conocimiento derivado de los hechos de
la experiencia ". En: ┐ Quķ es esa cosa llamada ciencia ?. pp.
11-25. SIGLO
XXI Editores S.A.
Curtis, H.; Barnes, N. 1993. "
La naturaleza de la ciencia ". En: BiologĒa, pp. 39-44. Ed. Med.
PANAMERICANA.
Dieterich, Heinz.
ōLa nueva guĒa para la investigaci¾n cientĒficaö. Ed. Planeta
Mexicana, S.A. de C.V., Mķxico,1996.
Kforoosh, Norma.
ōMetodologĒa de la investigaci¾nö. Noriega editores, Barcelona,
1998.
Eco, Humberto.
ōC¾mo se hace una tesisö. Ed. Gedisa, Espa±a, 1994.
Mardones, Josķ M.
ōFilosofĒa de las ciencias humanas y sociales. Materiales para una
fundamentaci¾n cientĒficaö. Ed. Prometeo, Mķxico,1999.
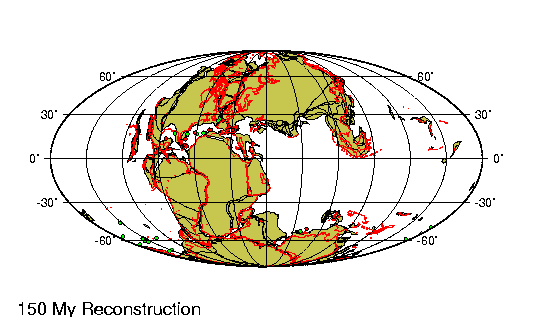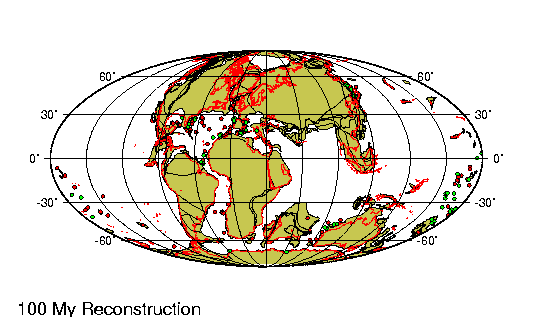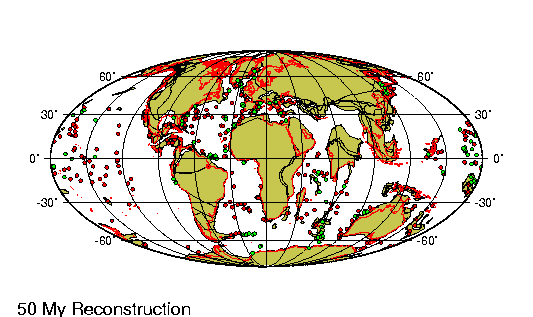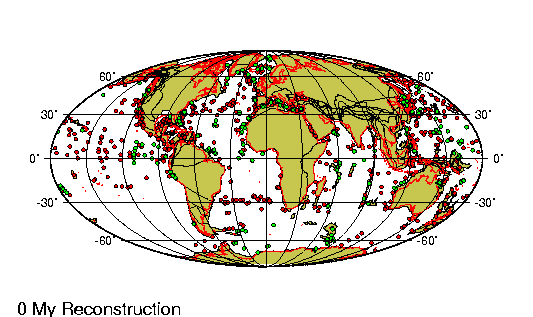
Origen de los continentes
y ocķanos.
|
Fragmento de: El origen de
los continentes y ocķanos. De Alfred Wegener. |
|
Traducci¾n de Francisco
Anguita Virella y Juan Carlos Herguera GarcĒa. |
La comparaci¾n de las estructuras
geol¾gicas a uno y otro lado del Atlßntico proporciona pruebas
rigurosas a nuestra teorĒa de que este ocķano representa una
descomunal fractura ensanchada cuyos bordes estuvieron en el pasado
en contacto directo o muy pr¾ximos. Esto es l¾gico, pues se podĒa
esperar que muchos pliegues y otras estructuras formadas antes de la
ruptura se continuasen a uno y otro lado, y realmente sus extremos
deben extenderse en ambos mßrgenes del ocķano, de forma que en la
reconstrucci¾n parecerĒan prolongaciones directas. Como esta
reconstrucci¾n viene forzada por el bien marcado trazado de las
lineaciones y no deja, por tanto, margen alguno para ajustes,
encontramos aquĒ un criterio totalmente independiente de la mayor
importancia para formarse un juicio sobre la correcci¾n de la teorĒa
movilista.
La fractura atlßntica presenta su
mayor anchura en el Sur, donde se form¾ primero: su anchura alcanza
aquĒ 6.220 km. Entre el cabo San Roque y el Camer·n hay s¾lo 4.880
km; a·n menos (s¾lo 2.410) entre el banco de Terranova y la
plataforma britßnica; entre el estrecho de Scoresby y Hammerfest
1.300, y entre el lĒmite Noroeste de la plataforma de Groenlandia y
las islas Spitzberg tan s¾lo de 200 a 300 km. En este ·ltimo punto
es donde la ruptura parece haber tenido lugar mßs recientemente.
Comencemos las comparaciones por el
Sur. En el extremo meridional de ┴frica se encuentra una cadena
plegada pķrmica (las monta±as Zwarten) que se alinea de Este a
Oeste. En la reconstrucci¾n, la prolongaci¾n hacia el Oeste de esta
cadena se encuentra justo al Sur de Buenos Aires, en una zona que no
presenta ning·n rasgo especial. Ahora bien, es muy interesante que
Keidel haya reconocido plegamientos antiguos en las sierras de esta
zona y especialmente en las meridionales, plegadas mßs intensamente;
plegamientos que por su estructura, su sucesi¾n litol¾gica y su
contenido f¾sil son totalmente similares no s¾lo a los de la zona
Noroeste de la Precordillera de las provincias de San Juan y
Mendoza, que se adaptan suavemente a los pliegues andinos, sino,
sobre todo, a los de las monta±as del Cabo, en Surßfrica. Afirma
este autor: ½En las sierras de la provincia de Buenos Aires,
especialmente en las cadenas meridionales, se encuentran series que
son muy parecidas a las de las monta±as del Cabo, en Surßfrica. Al
menos se da una gran coincidencia entre las tres formaciones
existentes: areniscas del Dev¾nico Inferior transgresivo en la base;
las pizarras fosilĒferas que marcan el punto de mßxima extensi¾n de
la transgresi¾n; y una formaci¾n superior muy caracterĒstica, el
conglomerado glaciar del Paleozoico Superior... Tanto los sedimentos
de la transgresi¾n dev¾nica como los conglomerados glaciares estßn,
al igual que en la cadena del Cabo, fuertemente plegados; y en ambas
zonas el movimiento principal estß dirigido hacia el Norte╗. De aquĒ
se sigue la conclusi¾n de que estamos ante un antiguo plegamiento
que pasaba por el extremo Sur de ┴frica y luego cruzaba Suramķrica
al Sur de Buenos Aires, para, finalmente, doblar hacia el Norte,
alineßndose con los Andes. Hoy, los fragmentos rotos de esa cadena
plegada estßn separados por un mar profundo de mßs de 6.000 km de
anchura. En nuestra reconstrucci¾n, que en este punto particular no
permite ning·n movimiento de ajuste, los fragmentos estßn en
contacto directo; sus distancias respectivas al cabo San Roque y al
Camer·n son iguales. Esta prueba de que nuestra composici¾n es
correcta resulta realmente espectacular y recuerda a las tarjetas de
visita rotas como se±al de reconocimiento. El que las monta±as Cedar
se separen de las cadenas surafricanas y se desvĒen hacia el Norte
en la costa es s¾lo un peque±o obstßculo a esta coincidencia, pues
esta rama casi extinguida muestra los caracteres de una desviaci¾n
local, que pudo ser causada por alguna discontinuidad en el lugar de
la fractura subsiguiente. Ramificaciones similares aparecen a·n en
mayor medida en las cordilleras europeas, tanto en las carbonĒferas
como en las terciarias, y no nos impiden sintetizar estas cadenas en
un sistema ·nico, y referirlas a una causa ·nica. Asimismo, aunque
(tal como se±alan los ·ltimos estudios) los plegamientos en ┴frica
hayan proseguido hasta tiempos mßs recientes, ello no implica que
existan diferencias de edad, pues, como leemos en Keidel: ½En las
sierras, la formaci¾n plegada mßs reciente es el conglomerado
glaciar; en las cadenas del Cabo, las pizarras de Ecca, en la base
de la Serie de Gondwana (series de Karroo) todavĒa muestran indicios
de plegamiento... En ambas zonas, los movimientos principales pueden
producirse en el intervalo entre el Pķrmico y el Cretßcico
Inferior╗.
Pero esta confirmaci¾n de nuestros
puntos de vista en la cadena del Cabo y su prolongaci¾n en las
sierras de Buenos Aires no es en absoluto un dato aislado, sino que
encontramos numerosas pruebas similares a lo largo de las costas
atlßnticas. Las gigantescas mesetas gnķisicas africanas, no
deformadas desde hace largo tiempo, muestran en sus grandes rasgos
una espectacular similitud con las brasile±as. Y esta similitud no
se limita a los caracteres generales, como lo demuestran por una
parte la coincidencia a una y otra parte de las rocas eruptivas y
los sedimentos, y por otra, la de las direcciones de los antiguos
plegamientos.
[...] En las direcciones de los
plegamientos primitivos que recorren estas grandes mesetas gnķisicas
encontramos otras concordancias. En el macizo gnķisico del
continente africano predominan dos direcciones (lineaciones) de
edades algo distintas. En el Sudßn domina la direcci¾n antigua,
orientada al Nordeste, que se evidencia en el trazado rectilĒneo del
curso superior del rĒo NĒger, y tambiķn en el Camer·n, y que corta
la costa con ßngulo pr¾ximo a los 45║; por el contrario, al Sur del
Camer·n pasa a primer plano la direcci¾n mßs moderna,
aproximadamente Norte-Sur, que discurre paralelamente a las curvas
de la costa.
En Brasil encontramos la misma
disposici¾n. Como escribe E. Suess: ½El mapa de la Guayana
oriental... muestra, en las rocas antiguas que componen esta zona,
direcciones aproximadamente Este-Oeste. Tambiķn las capas
paleozoicas que forman la parte Norte de la cuenca del Amazonas
siguen esta direcci¾n, y, por tanto, la lĒnea de costa que va desde
Cayena hacia la desembocadura del Amazonas es transversal a esta
direcci¾n... Por lo que se conoce actualmente de la estructura de
Brasil se puede aceptar que tambiķn hasta el cabo San Roque el
trazado de la costa es transversal a la direcci¾n de las monta±as,
pero a partir de este promontorio la lĒnea de costa viene marcada
por al direcci¾n de las monta±as, al menos hasta Uruguay╗. Tambiķn
en Amķrica los cursos fluviales (Amazonas por una parte, San
Francisco y Paranß por otra) siguen a grandes rasgos las lineaciones.
En realidad, los ·ltimos estudios, como, por ejemplo, el mapa
tect¾nico de Suramķrica formado por von Keidel siguiendo
esencialmente a J. W. Evans, han mostrado la existencia de una
tercera lineaci¾n paralela a la costa Nordeste, que complica
ligeramente las relaciones. Por ·ltimo, el considerable giro que
debe darse a Suramķrica en la reconstrucci¾n hace que la direcci¾n
del Amazonas se alinee paralelamente al curso alto del NĒger, de
modo que ambas lineaciones concuerdan con las africanas. AquĒ
podemos ver una vez mßs una confirmaci¾n directa de la antigua
conexi¾n entre ambos continentes.
[...] El conocido ge¾logo surafricano
Du Troit ha efectuado estudios comparativos particularmente
completos en un viaje de estudios a Suramķrica planeado con esta
finalidad. [...] Las coincidencias a ambas orillas del ocķano son
tan numerosas que actualmente ya no pueden considerarse
casualidades, sobre todo teniendo en cuenta que abarcan enormes
extensiones de terreno y un intervalo de tiempo que incluye desde el
pre-Dev¾nico hasta el Terciario. Du Toit a±ade: ½Por a±adidura,
ķstas que algunos llaman coincidencias son de naturaleza tanto
estratigrßfica como litol¾gica, paleontol¾gica, tect¾nica, volcßnica
o climßtica╗.
[...] ½A continuaci¾n vamos a
comparar las dos extensiones que van por una parte desde Sierra
Leona hasta El Cabo, y por la otra desde Parß hasta BahĒa Blanca,
limitßndonos en cada caso a una banda de unos 45║ de longitud y unos
10║ de latitud. En cada continente se puede observar:
╗1. El basamento estß compuesto de
rocas cristalinas de edad precßmbrica, con algunas capas de
sedimentos predev¾nicos, de edades variables aunque en general
indeterminadas: los caracteres litol¾gicos coinciden a grandes
rasgos.
╗2. En el extremo Norte de la zona
encontramos sedimentos marinos sil·ricos y dev¾nicos disconformes
sobre este complejo y muy ligeramente plegados, ocupando un amplio
sinclinal cuyo eje es oblicuo a la lĒnea de costa. Esta estructura
se extiende entre Sierra Leona y Costa de Oro en ┴frica y forma la
base del estuario del Amazonas en Suramķrica.
╗3. Hacia el Sur se encuentran, casi
paralelos a la costa, cinturones de rocas sedimentarias entre las
que predominan cuarcitas, pizarras y calizas. Sus edades son
Proterozoico y Paleozoico Inferior, y estßn plegados ligeramente en
el Norte y mßs intensamente en el Sur, donde son intruidos por masas
granĒticas, por ejemplo, en la zona entre L³deritz y Ciudad del Cabo
y entre el rĒo San Francisco y el RĒo de la Plata.
╗4. Al dev¾nico casi horizontal de
Clanwilliam corresponde una serie casi idķntica en Paranß y el Matto
Grosso.
╗5. A·n mßs hacia el Sur, el dev¾nico-carbonĒfero
de la parte meridional de la provincia del Cabo corresponde a la
comarca similar al Norte de BahĒa Blanca, incluyendo ambas una
transici¾n concordante hacia sedimentos glaciares pķrmicos y
carbonĒferos; las dos series estßn intensamente plegadas por
movimientos permotrißsicos y cretßcicos de direcciones similares.
╗6. Estas tillitas se contin·an hacia
el Norte, en ambos casos horizontales y transgresivas sobre el
dev¾nico, y descansando sobre una penillanura postglacial que se ha
formado en estas rocas y en otras anteriores y desaparece hacia el
Norte.
╗7. Los dep¾sitos glaciares estßn
cubiertos en ambos casos por sedimentos continentales pķrmicos y
trißsicos con flora de Glossopteris que cubren enormes extensiones y
van seguidos por potentes series de basaltos y doleritas a los que
se atribuye una edad Lißsico Inferior.
╗8. Estas capas de Gondwana se
extienden hacia el Norte del Karroo meridional hasta la zona de
Kaoko y desde Uruguay hasta Minas Geraes.
╗9. Afloramientos muy separados de
este mismo tipo se extienden tierra adentro hacia el Norte, en
Angola-Congo y en Piauh² y la zona del MaranhŃo.
╗10. Entre los sedimentos del
Trißsico Superior y del Pķrmico existe una interrupci¾n
intraformacional muy extendida, aunque en general no existe
discordancia angular, salvo en algunas zonas, donde los primeros
yacen en notable discordancia sobre los pķrmicos o antepķrmicos.
╗11. En la costa se encuentran
estratos calizos fuertemente inclinados, pero tan s¾lo en las
regiones de Benguela-Bajo Congo y BahĒa-Sergipe.
╗12. Sedimentos horizontales
cretßcicos y terciarios, tanto marinos como continentales, cubren
grandes extensiones entre Camer·n y Togo y en Cearß, MaranhŃo y
hacia el Sur, mientras que los extensos yacimientos en el Kalahari
pueden ser aproximadamente comparados con la formaci¾n Pampeana
ne¾gena y cuaternaria, en Argentina.
╗13. En este resumen general no puede
pasarse por alto el importante eslab¾n formado por las islas
Malvinas, cuyas series dev¾nico-carbonĒferas apenas se diferencian
de las de la provincia del Cabo, mientras que el Lafoniense es
totalmente comparable al sistema de Karroo. Tanto desde el punto de
vista estratigrßfico como estructural, las islas Malvinas tienen su
lugar junto a la regi¾n Suroeste del Cabo, y no en Patagonia.
╗14. Desde el punto de vista
paleontol¾gico, debemos fijarnos especialmente en: a) la ‘facies
austral’ del dev¾nico del Cabo, las islas Malvinas, Argentina,
Bolivia y el Sur de Brasil, opuesta a la ‘facies boreal’ del Norte
de Brasil y del Sahara central; b) el gķnero f¾sil Mesosaurus, un
reptil de la formaci¾n Dwyka, en El Cabo, y de las capas de Iraty,
en Brasil, Uruguay y Paraguay; c) la flora de Gangamopteris y
Glossopteris, con peque±as adiciones de formas septentrionales, en
las capas de Gondwana, al Sur de ambos continentes; d) la flora de
Thinnfeldia en la parte alta de esas mismas capas, en El Cabo y en
Argentina; e) la fauna neocomiense (uitenhaguiense) en el Sur de la
provincia del Cabo y en el Noroeste de Neuquķn, en Argentina; f) la
facies n¾rdica o mediterrßnea del Cretßcico y el Terciario al Norte
del Tr¾pico de Capricornio, y g) la facies suratlßntica-antßrtica
del Eoceno de Patagonia (formaci¾n San Jorge).
╗Por ·ltimo, y como punto n·mero 15,
tenemos los contornos geogrßficos de ┴frica y Suramķrica,
asombrosamente parecidos no s¾lo en general, sino tambiķn en los
detalles; y ademßs, excepto en el Norte, la orla de sedimentos
terciarios tiene peque±a extensi¾n, y por ello menor importancia.╗
[...] Du Troit considera, con raz¾n,
que el hecho de que las islas Malvinas, a pesar de elevarse en la
plataforma patag¾nica, no muestran ning·n parentesco con Patagonia y
sĒ con Surßfrica, constituye un apoyo especial para la teorĒa
movilista.
[...] Como ya se±al¾ antes, a base de
los datos paleontol¾gicos y biol¾gicos debe concluirse que los
intercambios de fauna y flora entre Suramķrica y ┴frica cesaron
entre el Cretßcico Inferior y Medio. No por ello se entra en
contradicci¾n con la suposici¾n de Passarge que la fractura entre
Surßfrica y Suramķrica ya estaba formada en el Jurßsico, pues la
ruptura se produjo gradualmente a partir del Sur, y, sobre todo, fue
precedida largo tiempo por la formaci¾n de fosas tect¾nicas.
En Patagonia, la ruptura tuvo como
consecuencia un movimiento continental peculiar, que A. Windhausen
describe del siguiente modo: ½La nueva revoluci¾n comenz¾ con
movimientos regionales de enorme escala, hacia la mitad del
Cretßcico╗, y realmente en este momento la llanura patag¾nica ½que
era una regi¾n de pendiente pronunciada, se convirti¾ en una
depresi¾n generalizada bajo condiciones ßridas o semißridas,
cubierta por desiertos de piedra y llanuras de arena╗.
Prosiguiendo hacia el Norte en la
comparaci¾n de las costas opuestas del Atlßntico, encontramos, en el
lĒmite Norte del continente africano, que las monta±as del Atlas,
cuyo plegamiento tuvo lugar principalmente en el Oligoceno, pero
habĒa comenzado ya en el Cretßcico, no encuentran continuaci¾n en el
lado americano.
Fuente: Wegener, Alfred.
El origen de los continentes y ocķanos. Traducci¾n de Francisco
Anguita Virella y Juan Carlos Herguera GarcĒa. Madrid: Ediciones
Pirßmide, S.A., 1983.
 Ver
mas ArtĒculos de Divulgaci¾n Paleontol¾gica Internacionales Ver
mas ArtĒculos de Divulgaci¾n Paleontol¾gica Internacionales |