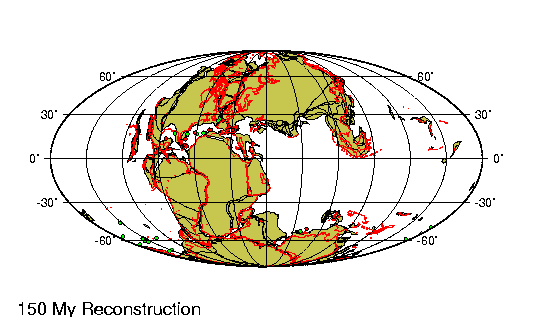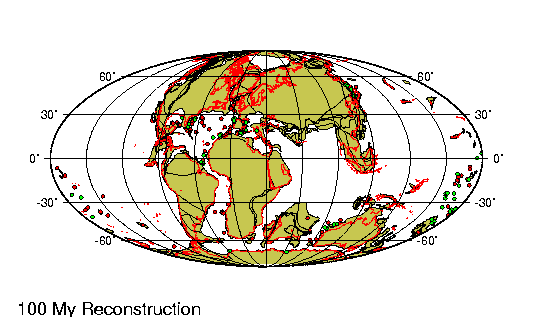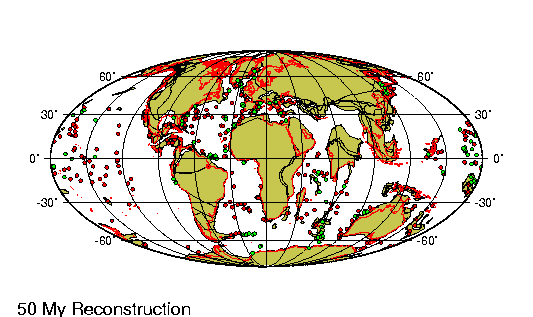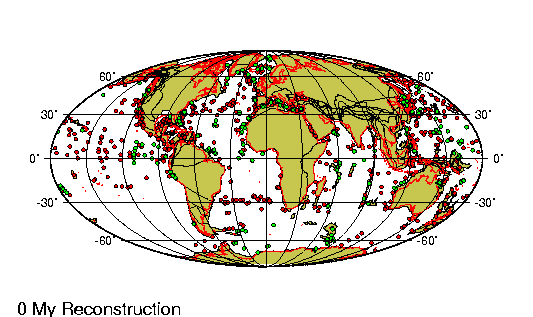
El Eoceno es la segunda
división del cenozoico, era de la escala de tiempos geológicos, que
comenzó hace unos 56,5 millones de ańos y finalizó hace unos 35,4
millones de ańos. Al igual que el paleoceno, que lo precedió, y el
oligoceno, que lo siguió, el eoceno (del griego eos 'alba' y kainos
'vida') fue definido en el siglo XIX por el geólogo británico
Charles Lyell sobre la base del porcentaje de especies modernas de
moluscos y crustáceos presentes en los estratos rocosos del
cenozoico.
En el hemisferio occidental, el eoceno marcó la última fase de la
orogénesis de las cordilleras, el episodio de alzamiento de las
grandes cadenas montańosas que se extienden hacia el norte y el sur
en el oeste de las Américas. En el noroeste, los sedimentos
arcillosos depositados en los vastos lagos del eoceno se compactaron
para formar valiosos depósitos de esquistos petroleros. Al mismo
tiempo, el supercontinente de Laurasia siguió desgajándose (véase
Tectónica de placas), la expansión de los suelos marinos comenzó de
hecho en la sección más septentrional de la dorsal Medioatlántica,
impulsando a Groenlandia hacia el oeste, alejándola del norte de
Europa, y desencadenando la erupción de grandes flujos basálticos,
cuyos restos pueden apreciarse en Irlanda, Escocia, Islandia y
Groenlandia.
Las fuerzas generadas por las colisiones continentales que habían
comenzado al principio de la era precedente, el mesozoico,
condujeron al alzamiento de los sistemas montańosos alpino e
himalayo. Mientras tanto, sobre las llanuras del noreste de la India
corrieron ingentes cantidades de basalto fundido al unirse este
subcontinente recién formado, desgajado de África durante el
cretácico, a Asia. En el hemisferio sur, la Antártida y Australia,
que habían estado unidas después de separarse de Gondwana en el
mesozoico, se separaron a su vez y se alejaron la una de la otra.
El clima del eoceno era subtropical y húmedo en toda América del
Norte. En las latitudes altas de Groenlandia y Siberia, en los
bosques templados húmedos, dominaban las secuoyas gigantes y los
árboles de hoja caduca como el haya, el castańo y el olmo. En el
clima por entonces cálido de Alaska, florecieron las higueras, las
magnolias y las cicadáceas. La rápida evolución de nuevos órdenes de
mamíferos, iniciada en el paleoceno, siguió adelante.
En Europa y Norteamérica aparecieron al
mismo tiempo formas ancestrales del caballo, el rinoceronte, el
camello y otros grupos modernos, como los murciélagos, los primates
y roedores similares a las ardillas. Muchos de ellos eran muy
pequeńos en comparación con las formas actuales. El caballo más
primitivo (Hyracotherium) medía unos 30 cm de altura y tenía tres
dedos en las patas traseras y cuatro en las delanteras. Los
carnívoros de aquel entonces, llamados creodontos, fueron el tronco
del que evolucionarían los perros y los gatos modernos. El final de
esta época fue testigo de la primera adaptación de los mamíferos a
la vida marina. En el norte de México, Egipto y Europa se han
encontrado restos fósiles de un animal similar a la ballena
pertenecientes al eoceno. El más grande de estos animales medía más
de 15 m, pero aún así era presa de los tiburones de entonces,
algunas de cuyas especies tenían mandíbulas de hasta 1,8 m de ancho.
América del Norte (Wyoming).
Tenia 45 cm de longitud. Con sus
patas cortas, las afiladas garras y la cola, larga y pesada,
es posible que Metacheiromys se pareciera a
una mangosta actual. Sin embargo, tenía la cabeza larga y
estrecha, como los armadillos. Poseía caninos fuertes, pero
había perdido casi todas las muelas. En su lugar, crecían
unas almohadillas córneas que el animal empleaba, sin duda
para triturar sus presas. El habitat de Metacheiromys
eran los bosques subtropicales densos que cubrían algunas
zonas del oeste de Estados Unidos durante el Eoceno. Las
garras de las patas delanteras eran mucho más grandes que
las ce las patas traseras, por lo cual es probable que Metacheiromys escarbara en busca de alimentos,
desenterrando hormigas, escarabajos y gusanos.
América del Norte (costa atlántica).
Tenia 6 m de longitud. Zygorhiza
pertenecía a una familia de ballenas primitivas que había
desarrollado un cuerpo muy alargado, como el de las
anguilas. La Zygorhiza propiamente dicha, sin
embargo, se parecía más a una ballena convencional que la
mayor parte de sus familiares. Su cuerpo abarcaba unas seis
veces la longitud del cráneo; esta proporción se repite en
la ballena actual. Pero, a diferencia de ésta, la cabeza se
unía con el cuerpo por medio de un cuello definido, aunque
corto, compuesto por el complemento de siete vértebras que
suele aparecer en los mamíferos. Las extremidades anteriores
tenían forma de aleta y es probable que se pudieran mover
desde el codo, a diferencia de las de las ballenas modernas,
cuyos miembros anteriores presentan los huesos fundidos y
rígidos hasta la parte superior del brazo. Es posible que
estas ballenas primitivas todavía se aparearan y se
reprodujeran en tierra, como sus antepasados anfibios. Las
articulaciones flexibles de sus miembros anteriores habrán
servido para halar sus cuerpos serpentinos fuera del agua,
sobre las rocas.
América del Norte (Wyoming).
Tenia 3 m de longitud; 1,5 m de altura en la
cruz. El grotesco Eobasilus se parecía
bastante a un rinoceronte con un par de colmillos caninos
con forma de sables en la mandíbula superior y seis
protuberancias óseas en la cabeza. Estos «cuernos» eran
romos y es probable que estuvieran recubiertos de piel.
También es probable que sólo el par situado en la parte
anterior de la nariz estuviera protegido por una vaina de
«cuerno» formada por una marańa de pelos, como en los
rinocerontes. Es posible que los machos los utilizaran para
embestirse de cabeza, en las luchas para determinar quién
era el jefe de la manada. Los incisivos de la mandíbula
inferior eran muy pequeńos y faltaban por completo en la
mandíbula superior, lo cual indica que la lengua y los
colmillos eran los órganos más importantes para reunir los
alimentos.
América del Norte (Wyoming).
Tenia 1,2 m de longitud. Visto desde una cierta distancia,
el cuerpo grueso, la cabeza corta y los pies planos de este
enorme animal deben de haberle dado el aspecto de un oso
moderno. Sin embargo, en cuanto abría la boca, sus enormes
incisivos cincelados le hacían parecerse más a una rata o un
conejo gigantescos. Igual que los de los roedores, los
incisivos estaban adaptados para roer: la superficie
anterior estaba recubierta de esmalte y crecía de forma
permanente durante toda la vida del animal. Los dientes para
moler, situados en la parte posterior de la mandíbula, se
gastaban continuamente, lo cual indica que Trogosus
comía sustancias vegetales abrasivas, tal vez raíces y
tubérculos que arrancaba de la tierra con las garras.
América del Norte. Los miácidos como
Vulpavus, fueron los primeros carnívoros auténticos;
aparecieron durante el Paleoceno, hace 60 millones de ańos.
Se trata de un grupo «artificial», ya que contiene animales
que no estaban muy relacionados entre sí. Sin embargo, al
clasificarlos es mejor separar estos carnívoros primitivos
de los más modernos. Los miácidos eran sobre todo mamíferos
pequeńos que vivían en zonas boscosas, donde no tenían
muchas probabilidades de fosilizarse. Los escasos restos que
se conservan radican que se parecían a los creodontos en
muchos aspectos, aunque es posible que fueran bastantes más
inteligentes y que sus dientes carniceros estuvieran mejor
desarrollados. Los huesos de las patas de algunos miacoideos,
como el Vulpavus del eoceno de América del
Norte, muestran que estos animales tenían patas muy móviles,
similares a las de los carnívoros modernos, que trepan a los
árboles en forma regular. El Vulpavus también
tenía garras curvas, que refuerzan esta idea. Sin embargo,
el estilo de vida trepador quizá no incluía a todos los
miacoideos. El Didymictis, del
paleoceno-eoceno de América del Norte y Europa,
probablemente vivía a nivel del suelo y habría sido un
corredor veloz o un cavador.
América del Norte. Los
dinocerados, mamíferos con "cuernos terribles", eran
criaturas herbívoras similares a los rinocerontes, famosas
por sus cuernos y colmillos. El dinocerado más primitivo, el
Prodinoceras, apareció en Asia durante el
paleoceno (65-53 millones de ańos atrás), pero casi todos
los tipos posteriores son de América del Norte. Cómo se
relacionan los dinocerados con los demás mamíferos aún es
tema de controversia. Probablemente hayan formado parte del
grupo de mamíferos con pezuńas(ungulados), ya que
presentaban similitudes con algunos mamíferos ungulados de
América del Sur. Otra posibilidad es que hayan estado
relacionados con los pantodontes y tilodontes. Una opinión
más controvertida es que los dinocerados descendieron de los
anagálidos, un pequeńo grupo de mamíferos similares a los
conejos. El dinocerado más grande y más conocido, el
Uintatherium, era tan grande como un rinoceronte
moderno. Se lo llamó así en 1872 por los indios Uintah, una
tribu que, como el Uintatherium, vivió en Utah.
Cuando se lo describió por primera vez, se discutió sobre si
el Uintatherium era o no un elefante. Hoy está
claro que los elefantes y los dinocerados no son parientes
cercanos. Las diversas formas que aparecían sobre los largos
cráneos de dinocerados como el Uintatherium y
el Eobasileus probablemente eran estructuras
de exhibición utilizadas para demostrar madurez sexual. Los
machos parecen haber tenido cuernos más grandes y una
proyección mayor en la mandíbula. Es probable que lucharan
usando estas estructuras, quizá empujándose unos a otros con
los cuernos y crestas y mordiéndose con los colmillos.
Utah (EU); Mongolia. Los
uintateres fueron grandes herbívoros que vivieron durante
los primeros 30 millones de ańos de la era de los mamíferos
en América del Norte y Asia Central. El Dinoceras,
un uintatere característico, era grande como un rinoceronte,
pero no se emparentaba con las formas modernas. La cabeza
estaba adornada por tres pares de protuberancias óseas, dos
sobre las fosas nasales, dos sobre los ojos y dos en la
parte posterior. Además, los machos poseían un par de
colmillos largos como cuchillos. Tales protuberancias y
colmillos tal vez se usaron en las luchas entre los machos
por las hembras o para defenderse, aunque había pocos
carnívoros en esa época que pudieran enfrentarse al
Dinoceras.
América del Norte.
Los cánidos, que incluyen
los zorros, chacales, coyotes, lobos y perros actuales, son
un grupo próspero y versátil. Cuentan con una historia
evolutiva de unos 40 millones de ańos, y se han adaptado a
una gran diversidad de habitas y a una amplia variedad de
dietas. Como miembros del orden de los carnívoros, están
relacionados con Ias nutrias y las comadrejas, los felinos y
las mangostas, y con las focas, los leones marinos y las
morsas. Los cánidos más primitivos, conocidos por primera
vez a fines del Eoceno, hace unos 40 millones de ańos, eran
unos animales de patas relativamente cortas, que se
asemejaban a las mangostas y las civetas, más que a los
perros. Estaban confinados a América del Norte, el centro de
la evolución de los cánidos, de forma casi exclusiva: la
familia no colonizó otros continentes hasta el final del
.Mioceno, hace apenas 6 millones de ańos.De los cinco
géneros que existían a principios del Oligoceno (35 millones
de ańos atrás), al finalizar el Mioceno (entre 10 y 6
millones de ańos atrás) los cánidos se habían diversificado
hasta formar 42 géneros, y desde entonces han ido decayendo
hasta los doce géneros que sobreviven en la actualidad,
entre los que se incluyen los perros domésticos. El
Hyaenodon era un animal con hábitos depredadores,
semejantes a los de un lobo. Forma parte de un grupo llamado
creodontes, que no son considerados verdaderos carnívoros.
Los creodontes vivieron desde el paleoceno hasta el mioceno,
y se habrían parecido a las civetas, los gatos o los perros
modernos. Los dientes cortantes ubicados en la parte
posterior de las quijadas muestran que estos animales eran
depredadores.
Perú. Medía
1.5m. y pesaba 54 kilos. Vivió en el Eoceno tardío, hace 36
millones de ańos, un periodo caracterizado por el clima
cálido y una fauna rica en lo que hoy es la Reserva Nacional
de la Bahía de Paracas: colosales tiburones, variados
cocodrilos, cetáceos primitivos y aves gigantescas. El
artículo de la revista Science trata de esta investigación
que se centró sobre todo en las plumas del animal. La
fosilización de partes blandas es muy difícil; pero en este
caso se han conservado no solo las plumas del cuerpo y las
alas, sino las escamas de las patas. Por primera vez se ha
revelado la forma de las plumas de los pingüinos
prehistóricos y hasta el color. El Inkayacu
paracasensis era gris y marrón rojizo, a diferencia
de los pingüinos modernos que son negros y blancos. La
importancia de la conservación de las plumas permite
estudiar la evolución de los especímenes de esa época hasta
el diseńo de las plumas de los pingüinos actuales.
Pelagornithidae.Fürbringer,
1888.
Húmero de este ejemplar de
dimensiones extraordinarias. Prensa.
Esqueleto de
un pelagornítido, en el Museo Argentino de Ciencias
Naturales de Buenos Aires.
Aspecto del pelicano
gigante de la antártica. Prensa.
Antártica,
Argentina.
Familia extinta de grandes aves marinas. Sus restos fósiles
se han hallado en muchas partes del mundo en rocas que datan de
finales del Paleoceno hasta el límite entre el Plioceno y el
Pleistoceno.
Superaba cómodamente los
seis metros de extensión con sus alas abiertas. Era un ave
extremadamente ligera para su tamańo, casi como una pluma, que solo
pesaba unos 30 o 35 kilos. Podía recorrer grandes distancias sobre
los mares y cazaba peces durante vuelos rasantes. Sus restos fueron
encontrados por paleontólogos argentinos cerca de la base Marambio. En la Antártida existieron dos grupos de
pelagornítidos: uno de ellos estaba compuesto por aves que no
superaban los 5 metros de envergadura alar, mientras el otro tenía
representantes gigantes que podían alcanzar entre seis y siete
metros. Hay evidencias de que, hace 50 millones de ańos, se inició
un período de calentamiento de la temperatura de los océanos, el
cual provocó seguramente una gran productividad biológica de los
mares antárticos y permitió que los pelagornítidos y los pingüinos
tuvieran alimento suficiente para poder desarrollar tamańos tan
gigantescos. Para sujetar su alimento, los pelagornítidos tenían
unos pseudodientes. Se trataba de unas expansiones óseas en sus
picos, pero no tenían la capacidad de mordida de aquellos pingüinos
gigantes con los que convivieron, ya sus huesos del rostro no
estaban preparados para tener mucha resistencia; posiblemente,
tenían una alimentación parecida a la de un pelícano actual, que se
abastece de animales blandos, como calamares o peces. Había allí un
ambiente costero, poblado de muchas especies de pingüinos y
gaviotas, y muy próxima a esa costa había un ambiente boscoso
habitado por comadrejitas, marsupiales del tamańo de un ratón,
ungulados ya extintos del tamańo de una oveja y allí también
encontramos hace poco al falcónido más antiguo del mundo. Los
doctores Marcos Cenizo, Carolina Acosta Hospitaleche y Marcelo
Reguero son los autores del estudio.
Fragmento craneal de
Paraptenodytes
antarticus.(*)
Fósiles de
Paraptenodytes
antarticus,
en el
Geoparque Bryn Gwyn de
Gaiman, Chubut.
Aspecto del pingüino fósil
Paraptenodytes. (*)
Patagonia,
Argentina.El género Paraptenodytes fue acuńado por
Florentino Ameghino en 1891 para designar a los especímenes fósiles
originalmente descritos por François Moreno y Alcide Mercerat como
Palaeospheniscus antarcticus. La especie tipo del género
corresponde a Paraptenodytes antarcticus, cuyo
holotipo se encuentra depositado en el Museo de La Plata, proceden
de la Formación Monte León. En 1946, George Gaylord Simpson
describió un nuevo espécimen de la especie tipo, el cual incluía
parte del cráneo y postcráneo asociados. Este hallazgo fue durante
ańos el esqueleto de pingüino fósil más completo conocido y en el
único con cráneo asociado hasta la descripción de
Marplesiornis en 1960. Paraptenodytes
era
un verdadero genero de pingüino prehistórico de hábitos acuáticos
como los actuales, pero algunas de sus especies tenían un gran tamańo,
ya que oscilaban desde 0,20 a 1,90 metros de alto. Durante este
mismo Periodo, vivió en la Patagonia otro genero de pingüino también
desaparecido, cuyo tamańo era similar a sus parientes actuales,
llamado Apterodytes, caracterizándose por tener unas alas ridículamente
pequeńas. Su comportamiento no era muy distinto a sus
descendientes, ya que anidaban en las costas marinas, donde cavaban
su propio nido o aprovechaban aquellos abandonados por otros
animales, como marsupiales y reptiles. Su alimentación estaba
constituida principalmente de peces de tamańo variante y de
numerosos invertebrados, los cuales se hallan normalmente asociados
a estos vertebrados emplumados. El enigma de estas aves
desaparecidas es si para este periodo habían desarrollada la glándula
uropigia, la cual se encargaría de distribuir aceite natural por su
cuerpo, permitiendo la impermeabilización de las plumas y evitando
de que estas presenten hipotermia en aguas frías. Recordemos que en
este Periodo Patagonia era una selva Sub-tropical con temperaturas
constantes de unos 20° centígrados durante gran parte del ańo. Los
principales restos de Paraptenodytes proceden de la
Formación Sarmiento y Gaiman.
Antarctoboenus carlinii.
Cenizo,
Noriega y Reguero, 2016.
Resto fósil antártico de
Antarctoboenus
carlinii. Imagen Fundación Azara, prensa.
Posible aspecto del primitivo
Halcón del Eoceno del sector antártico Argentino.(*)
Antartica, Argentina.
Los
investigadores Marcos Cenizo (Museo de
Historia Natural de La Pampa - Fundación
Azara), Jorge Noriega (CICYTTP - CONICET -
UADER) y Marcelo Reguero (Instituto
Antártico Argentino) dieron a conocer un
falcónido ancestral que vivió hace 50
millones de ańos atrás en lo que hoy es la
isla Marambio, en el sector antártico
argentino. El nuevo “protofalcónido”
-al que los autores denominaron,
Antarctoboenus carlinii- representa
el miembro más antiguo de este linaje de
rapaces, duplicando en edad a los restos de
mayor antigüedad que se conocían hasta el
momento. Este descubrimiento confirma
que los falcónidos se originaron en algún
lugar del hemisferio sur entre América del
Sur y la Península Antártica, algo que ya
anticipaban los estudios evolutivos basados
en datos genéticos obtenidos de las especies
vivientes. La edad del Antarctoboenus
carliniiofrece un nuevo punto de
calibración a los relojes moleculares que
permiten inferir los momentos del tiempo
geológico en que los principales linajes de
aves divergieron unos de otros. El nombre
genérico del nuevo espécimen significa
“caminante antártico” y fue nominando en
honor al querido y siempre recordado Dr.
Alejandro Carlini, líder por casi 25 ańos
del Programa de Mamíferos Marinos del
Instituto Antártico Argentino y uno de los
investigadores argentinos más destacados en
el conocimiento de los ecosistemas
antárticos.
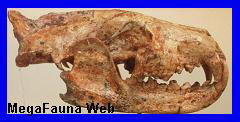
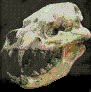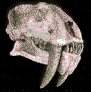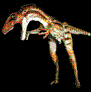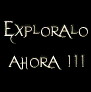
Niolamia argentina.Ameghino, 1899.
Cráneo de Niolamia
de unos 30 cm de ancho, con grandes cuernos laterales, en el
Museo de La Plata. Placas
tentativamente atribuidas a Niolamia (tortuga gigante de
tierra).
Reconstrucción del aspecto de Niolamia
argentina (*).
Patagonia,
Argentina.
Es una de las tantas especies que sobrevivieron al cataclismo que
extinguió a los dinosaurios. Fue
una tortuga terrestre de tamańo colosal y comparable a los grandes
gliptodontes del Pleistoceno. Su cabeza estaba armada por
grandes protuberancias, dos de las cuales se prolongan de manera
extraordinaria, semejantes a los cuernos de un buey. La cola no
pasaba de percibida, ya que se encontraba protegida por un estuche
óseo compuestos por varios anillos imbricados y con protuberancias
cónicas. Los primeros restos de esta tortuga fueron hallados en la
Provincia de Neuquén en el Cretácico superior. Florentino Ameghino
lo dio a conocer en 1899 y lo bautizo con el nombre de
Niolamia, para diferenciarla de la gran tortuga australiana
del Pleistoceno conocida como Meiolania, con la cual
Ameghino creía ligeramente relacionada. Llegaron a medir hasta 2,5
metros de largo. En América del Sur esta especie se prolongo hasta
el Eoceno.
Madtsoia
bai. Simpson,
1933
Restos fósiles constituido por una gran cantidad
de vértebras y costillas articuladas. Cráneo muy completo,
encontrados en la Patagonia Argentina.
"American
Museum of Natural History" in New York. Al lado
reconstrucción de Madtsoia. (*)
Patagonia, Argentina.En la actual provincia de Chubut, al sur de la republica Argentina,
durante el Eoceno, el territorio era muy distinto al desierto que
hoy podemos observar. Para entonces el clima era sub-tropical con
alta humedad. Así lo indica la presencia de primitivas tortugas y
cocodrilos. También el registro paleobotánica, donde se han podido
identificar grandes palmeras y otras plantas de zonas lluviosas. Uno
de los primeros registros de grandes serpientes del Terciario de
Patagonia fue publicado por el sabio Florentino Ameghino en 1906.
Madtsoia, fue un enorme "ofidio" (víboras y serpientes)
hallado en el ańo 1933 por el paleontólogo George Gaylord
Simpson. Los fósiles hallados, corresponden a un esqueleto
incompleto, pero con una excelente conservación. Se trata de una
columna vertebral articulada y con sus respectivas costillas. Los
estudios revelaron que este enorme animal poseía mas de 300 o 400 vértebras,
y que en vida superaba los 10 metros de largo. (las Anacondas miden
8 metros). Aun se discute si fue una serpiente de hábitos
terrestres o acuáticos, pero si estamos seguros que se alimentaría
de mamíferos notoungulados de tamańo medio y chico, a los cuales acechaba
para enrollarse en su cuerpo y asfixiarlos hasta poder digerirlos.
En 1959 se dio a conocer la existencia de una rama mandibular de
gran talla, la cual fue hallada en en el Paleoceno tardío en la
localidad de Gaiman, al norte del Rió Chubut, atribuida al genero
Madtsoia, cuya antigüedad seria entre 55 y 60 millones de ańos.
Los primeros restos y que corresponden a este Periodo (Eoceno
temprano), corresponden a la localidad de Cańadon Vaca, en el
Sudoeste de la Provincia de Chubut, Argentina. Durante el Mioceno
vivieron otros "Ofidios" de gran tamańo (ver).
Fragmento de vértebra de Boidedae,
comparada con un representante actual (en su interior).
(*)
Boidedae
capturando un Astrapotherium.
(*)
Patagonia,
Argentina.Los boidos o boas (Boidae) son una familia de serpientes
constrictoras, es decir que matan a sus presas por constricción,
encerrándolas en sus anillos. En
1986, mientras que la Paleontóloga Adriana Albino consultaba la
colección del Museo de La Plata, descubrió una vértebra de
serpiente y de tamańo gigantesco, tanto que figuraba en el registro
del Museo como perteneciente a un cocodrilo prehistórico. El
estudio de la misma dio como resultado que no pertenecía al genero
descrito mas arriba (Madtsoia), sino un representante fósil de los
grandes Boideos actuales. Así mismo la comparación del fósil con
restos de una Boa, dio como resultado de que se trataba de un animal
joven, que aun no alcanzo su máximo desarrollo corporal. El numero
de vértebras de estos animales varia entre las 300 y 400
aproximadamente. Asumiendo que las formas fósiles se mantuvieron en
dicho rango de variabilidad, estimamos, en base a la longitud
corporal de la forma juvenil habría sido entre los 5 y 7 metros,
mientras que los adultos tendían entre 10 y 12 metros, es decir,
algo mas grande que lo calculado para Madtsoia. Es muy probable que
capturaba a sus presas cuando estas se encontraban bebiendo agua a
las orillas de algún espejo de agua o bien cuando estas estarían
descansando, momento de mucha vulnerabilidad, que estos enormes
Ofideos aprovecharían para alimentarse, o bien, a presas muy
adultas o enfermas. Sus restos proceden de los niveles del Eoceno
temprano de la zona de Valle Hermoso, al Sudoeste de la Provincia de
Chubut.
Llanquibatrachus truebae.
Baez y Pugener 2003.
Rana fósil - Confluencia (Neuquén) del Eoceno,
Terciario Museo
Paleontológico de Bariloche (Río Negro)
Recreación de un
anfibio pípido. (*).
Patagonia, Argentina.Se
trata de un anfibio pípido único en su tipo, ya que se encontraron
fósiles de todas sus etapas larvales (renacuajos), incluyendo sus partes
blandas. Fue hallado frente al Lago Nahuel Huapi, en la ciudad de
Bariloche. La anatomía de su
mano indica que se alimentaba por partículas pequeńas en suspensión. La
distribución actual disyuntiva de este grupo de anfibios en América del Sur
y África demuestra las conexiones de la fauna dulceacuícola entre ambos
continentes (ya que los anfibios no soportan aguas marinas). Su característica básica es que carecen de lengua, por
lo que se les llama también Aglosos. La pipa o rana de Surinam en la
actualidad habita en
las regiones tropicales sudamericanas; es de costumbres acuáticas y
cuerpo aplastado; los ojos son muy pequeńos, y las crías no pasan por la
fase larvaria acuática, sino que la viven dentro del huevo. Otro anfibio
conocido; Shelania pascuali, que precede de la Laguna del
Hunco, en la provincia de Chubut.
Callistoe vincei.Babot,
Powell & Muizon, 2002.
Mariano Magnussen
Saffer, junto al fósil de Callistoe
vincei,
en el Museo de Ciencias Naturales "Miguel Angel Arra" de la
Universidad Nacional de Salta.
Reconstrucción paleoartistica
del primitivo
Callistoe
vincei,
del Eoceno de la Provincia de Salta, Argentina. (*).
Norte,
Argentina.Fue un interesante Proborhyaenidae, una familia extinta de
marsupiales del orden Sparassodonta, que vivieron en Sudamérica desde el
Eoceno (Casamayorense) hasta el Oligoceno (Deseadense). En ocasiones han
sido incluidos como subfamilia de sus parientes los borhiénidos (Borhyaenidae).
Fueron metaterios carnívoros, usualmente de gran tamańo, estando entre
los mayores representantes de los esparasodontes y de los metaterios
carnívoros en general. Los proborhiénidos se caracterizaban por un
cráneo grande y macizo, con molares especializados en el consumo de
carne (carnasiales)y sus colmillos carecían de esmalte dental en los
ejemplares conservados; debió de ser muy delgado o restringido a las
puntas en vida. En el género Aminiheringia los colmillos
están además orientados hacia adelante. Estos dientes, igual que en los
tilacosmílidos presentaban una raíz abierta incluso de adultos, con lo
que estos colmillos podían crecer continuamente. Esta característica fue
sugerida como evidencia de su parentesco cercano con Thylacosmilus,
pero dicha hipótesis ha sido descartada.Callistoevincei,
un fósil muy completo, procede de la Formación Lumbreras,
en la localidad de Pampa Grande, Provincia de Salta. Fue el equivalente
ecológico de los grandes canidos de América del Norte. De cráneo y
mandíbulas alargadas, con un robusto arco cigomatico y una poderosa
musculatura masticatoria, típico del habito depredador. Seguramente se
alimento de mamíferos notoungulados y armadillos, comunes en su época.
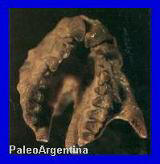
Notostylops
brachycephalus. Ameghino, 1904.
Esqueleto de Periphragnis
(semejante a Nototylops)
hallado en La Gran Hondonada,
en el MEF. Museo Egidio Feruglio de Trelew.
Cráneo con
detalles del maxilar de Notostylops del Eoceno inferior
de Patagonia. Museo de La Plata.
Reconstrucción de Notostylops.
(*)
Patagonia, Argentina. Su
nombre significa "columnas traseras". Pertenece a un grupo
único de animales del orden "Notoungulados" y que no se
pueden hallar en otras partes del mundo porque evolucionaron en el
largo aislamiento del continente Sudamericano. Uno de los miembros
mas primitivo de este grupo fue Notostylops, que tenia un tamańo
algo chico (75 centímetros) a diferencia de sus parientes del
Plioceno y Pleistoceno. Poseían unos dientes enormes muy parecidos
al de los roedores, con un frente largo y filoso para cortar hojas,
romper frutos duros y plantas herbáceas. Una de las características
mas importantes del cráneo de este animal y que luego se repetiría
durante los otros periodos, fue lo pronunciado en su oído medio, lo
que sugiere podía escuchar desde grandes distancias.
Propalaehophophorus australis.
Parte de
coraza y cráneo de Propalaehophophorus en el MEF.
Coraza de Propalaehophophorus.
(*)
Reconstrucción en vivo del primitivo Propalaehophophorus australis.
(*)
Patagonia, Argentina. Este
genero pudo ser el antecesor de los Gliptodontes, cuyas características
principales se encuentras desarrolladas en el Pleistoceno de la
presente PaleoGuia (ver). Algunas de las características propias de
este antiguo representante del Eoceno medio de la Patagonia
Argentina, se basa principalmente en poseer aun la cola como los
Dasipodidos vivientes (mulitas y peludos), sin escudetes y tubos óseos.
Estas estaban formadas por escamas y placas indicadas y libres. La
coraza dorsal conserva a sus lados inferiores grandes hendiduras
verticales que separaban a las bandas transversales con cierta
flexibilidad, las cuales se podrían interpretar como los últimos
vestigios de las bandas móviles de los armadillos. El tamańo de
este genero, era poco mayor al Tatu Carreta actual, el cual aun
habita el Noroeste Argentino.
Catogenus punctatus.
Fósil de Catogenus
punctatus del Museo Dr. Rosendo Pascual del lago
Gutiérrez en Bariloche. Imagen de prensa.
Aspecto de un Passandridae.
Imagen ilustrativa. (*).
Patagonia, Argentina.Se dio a conocer el hallazgo de un insecto
fosilizado, un escarabajo de 11,75 milímetros único en el mundo, cerca
de Pilcaniyeu (provincia de Río Negro), que tiene una antigüedad de 47,5
millones de ańos. Catogenus punctatus sp. nov. es la
primera especie fósil de la familia Passandridae (Coleoptera: Cucujoidea)
descripta en el mundo. Su descubridor es el paleontólogo Julián
Corsolini, director del Museo Dr. Rosendo Pascual del lago Gutiérrez en
Bariloche, y la investigación de sus características más la publicación
del trabajo científico, la realizó junto al biólogo Leonardo Ramírez del
Centro Austral de Investigaciones Científicas de Ushuaia, y al biólogo
recientemente fallecido Osvaldo Di Iorio de la Facultad de Ciencias
Exactas y Naturales de la Universidad de Buenos Aires. Es la primera vez
que se encuentra en el mundo esta especie y fue encontrado en el mismo
lugar donde se halló la flor más vieja del mundo. En el mismo estrato se
encontraron hojas y arańas fosilizadas. El material procede de
Pilcaniyeu, de la estancia Don Hipólito, a unos 60 km de Bariloche. Con
respecto a la paleoecología o paleoambiente, tanto los insectos como las
hojas y la interacción entre ellos nos puede decir mucho, en ese caso el
ambiente tenía una temperatura entre 19 y 25 grados, era subtropical, y
eso nos puede dar pautas de dónde estaba el Ecuador, si estaba donde
está hoy o más abajo, cómo estaban los polos.
Anisoptera.
Selys, 1854.
Larvas de libélulas fósiles,
del Eoceno de la zona del Río Pichileufu. Museo Paleontologico
de Bariloche.
Posible aspecto de las
libélulas del Eoceno patagonico. (*).
Patagonia, Agentina.
Durante el Eoceno de la Patagonia Argentina se observa una gran variedad
de insectos, los cuales llegan a nuestros días convertidos en fósiles.
Entre ellos podemos encontrar larvas del Infraorden Anisoptera, una
especie de libélula que vivió en la actual zona del Río Pichileufu,
próximo a la ciudad de San Carlos de Bariloche, donde el Museo
Paleontológico de esa ciudad a colectado una gran muestra de distintos
insectos y otros invertebrados. Los Anisoptera son paleópteros, es
decir, insectos que no pueden plegar las alas sobre el abdomen. Se
caracterizan por sus grandes ojos multifacetados, sus dos pares de
fuertes alas transparentes y por su abdomen alargado. Se alimentan de
mosquitos y otros pequeńos insectos como moscas, abejas, mariposas y
polillas. Su hábitat natural se encuentra en las cercanías de lagos,
charcos, ríos y tierras pantanosas, ya que sus ninfas son acuáticas.
América del Norte.
El
primate de 47 millones de ańos, Notharctus tenebrosus,
claramente tenía una garra de aseo en su segundo dígito
-sorprendentemente, la garra era algo aplastada, como un
clavo. Según el investigador, Notharctus puede
proporcionar evidencia de que las uńas se desarrollan a
partir de este grupo de primates, o que, por otro lado, las
garras se desarrollaron a partir de las uńas en este grupo.
La presencia o ausencia de una garra de aseo se ha utilizado
anteriormente para clasificar a los grupos de primates: los
seres humanos, los simios y los monos tienen uńas, mientras
que los lémures tienen garras de aseo en el segundo dígito.
Ademáslas uńas pudieron ser el punto de partida, y las
garras de aseo se desarrollaron a partir de éstas como un
rasgo funcional.
África y Asia (zona mediterránea). Tenia
2,5 metros longitud. Protocetus, que vivió apenas unos 8 millones de ańos después
que Pakicetus, había adquirido un aspecto mucho más
semejante al de la ballena. Su cuerpo era más fusiforme,
aproximándose a la forma de las ballenas actuales. Las
extremidades anteriores eran planas y con
forma de aletas, pero las posteriores estaban muy reducidas,
y si bien es posible que todavía sobresalieran del cuerpo,
no habrán servido de mucho para nadar.
Es probable que hubieran aparecido un par de lóbulos
horizontales, llamados aletas, en la cola de Protocetus, a
juzgar por la estructura de las vértebras en esta zona. Su
movimiento hacia arriba y hacia abajo proporcionaba la
fuerza propulsora necesaria para conducir al animal a través
del agua.
El cráneo de Protocetus se había vuelto bastante largo, con
un hocico estrecho. Los dientes eran puntiagudos y estaban
dispuestos en zigzag en la parte anterior de las mandíbulas.
Estos dientes retenían a la presa, mientras los posteriores
la cortaban en trozos. Es indudable que Protocetus y otras
ballenas primitivas buscaban peces en las aguas costeras
poco profundas. Las fosas nasales de esta ballena habían
comenzado a retroceder en su cabeza, alejándose de la
posición que ocupaban en las ballenas más primitivas, .en el
extremo del hocico. Protocetus todavía poseía un olfato
agudo, pero es probable que la visión fuera el sentido que
más utilizara para cazar a sus presas. En contraste con Pakicetus, sus orejas estaban adaptadas para oír debajo del
agua, pero es poco probable que ya hubiese desarrollado el
sistema de detección por ultrasonidos que utilizan las
ballenas.
Frankfurt, Alemania.Con
un tamańo aproximado al de un pequeńo gato, el animal tiene
cuatro patas y una larga cola. Nadie sostiene que es un
ancestro directo de monos y humanos, pero ofrece algunos
rasgos de cómo podría haber sido un antiguo ancestro,
dijeron los investigadores. Un equipo de científicos ha
anunciado el descubrimiento del esqueleto fosilizado de un
mono-lemur de 47 millones de ańos de antigüedad que podría
convertirse en el eslabón perdido de la evolución humana. El
fósil, al que han llamado “Ida” ha sido presentado en una
rueda de prensa especial en Nueva York. El descubrimiento
del 95% del esqueleto de un mono-lemur ha sido descrito por
los expertos como "la octava maravilla del mundo", y
consideran que su impacto en el mundo de la paleontología
será algo así como "un asteroide caído en la Tierra" al
considerar que se ha que inició Charles Darwin hace 200 ańos
con la Teoría de la Evolución. Así, el equipo investigador
seńala a Ida como la prueba de la transición de las especies
que inició Darwin a bordo del Beagle. Según expresó sir
David Attenborough al canal de televisión Sky News, Darwin
"se habría emocionado" de haber visto el fósil porque
expresa lo que el ser humano es y de dónde procede. De
acuerdo con los estudios radiométricos de las rocas
volcánicas de Messel, los expertos descubrieron que Ida
vivió hace 47 millones de ańos en el periodo del Eoceno,
cuando los bosques tropicales casi se juntaban con los polos
y Suramérica estaba aún separado de Norteamérica.
Inglaterra; Wyoming (EU);Asia. La historia de la
evolución de los caballos se conoce bien. El primer caballo
fue el Hyracotherium, un animal del tamańo de
un perro terrier que probablemente no se parecía mucho a un
caballo. Tenía patas cortas con cuatro dedos y un cráneo
parecido al del perro con dientes cortos. Probablemente
llevaba una vida secreta en la maleza de los tupidos bosques
subtropicales que cubrían América del Norte y Europa hace 50
millones de ańos, alimentándose con hojas y brotes. Los
primeros fósiles de Hyracotherium se hallaron
hace 140 ańos en Inglaterra. Más tarde, algunos restos de
caballos primitivos se encontraron en América del Norte y se
les llamó Eohippus, "caballo del amanecer".
Tiempo después quedó claro que el Eohippus era
el mismo Hyracotherium europeo. Al mismo
tiempo, se encontró una serie de fósiles de caballos que
parecen ligar al pequeńo Hyracotherium del
bosque con los grandes caballos modernos de las praderas.
Alemania.
Los pangelines, que tienen escamas y se alimentan de
hormigas, son los mamíferos modernos más peculiares y raros.
Hay siete especies vivas en África y el sureste de Asia,
pero hay pocos rastros fósiles. Un hallazgo notable se hizo
en Alemania en 1978: el Eomanis, el más
antiguo fósil de pangelín conocido. El hallazgo fue
inesperado, pues los pangelines modernos habitan muy lejos
de Alemania. El Eomanis tenía un largo cráneo
en forma de tubo y sin dientes, así como una débil mandíbula
inferior que no servía de mucho. Como los pangelines
modernos, el Eomanis pudo poseer una larga y
musculosa lengua que servía para atrapar docenas de hormigas
a la vez. Se conoce la dieta exacta del Eomanis
porque los restos de su última comida se conservaron en la
región del estómago en el esqueleto fósil. Había tanto
restos de insectos como fragmentos de plantas. Eomanis
muestra las otras características de los pangelines. Las
patas son cortas y están equipadas con largas garras que
sirven para cavar en los hormigueros y termiteros. Además,
el cuerpo estaba cubierto de anchas escamas sobrepuestas de
un material parecido al de nuestras uńas. Sin duda, el Eomanis podía enroscarse y formar una bola
blindada cuando era amenazado, tal como hacen sus parientes
modernos.
Eurotamandua joresi.
Storch and Habersetzer, 1991.
Europa (Alemania).
Tenia 90 cm de longitud. Hasta hace muy poco, cuando se
descubrió un oso hormiguero fósil en los depósitos de
esquisto petrolífero situados cerca de Frankfurt, en
Alemania, y se le bautizó con el nombre de
Eurotamandua. los paleontólogos pensaban que sólo
había habido osos hormigueros en América del Sur. Por su
hocico largo y tubular, las mandíbulas débiles y sin
dientes, y los fuertes miembros delanteros provistos de
garras enormes, no cabía duda de que se trataba de un oso
hormiguero, y parece que era muy similar al oso hormiguero
con cuello. Tamanduá. El registro fósil de los
osos hormigueros es bastante pobre, de modo que no se sabe
cómo encaja este descubrimiento europeo dentro de la
historia evolutiva general del grupo. Existen pruebas de la
alimentación típica de los osos hormigueros que seguía Eurotamandua por las hormigas fosilizadas que se
encontraron en el emplazamiento alemán.
Europa (Alemania). Tenia 75 cm de longitud. Es
probable que Leptictidium se pareciera a las
musarańas elefantes actuales, salvo por las patas traseras y
la cola, que eran más largas. Fue un corredor bípedo, como
los seres humanos y algunos de los dinosaurios carnívoros
más pequeńos. Las patas traseras eran largas, ligeras y
parecidas a las de las aves, y la mayor parte de los
músculos se concentraban en torno a los muslos. Las
extremidades anteriores medían menos de la mitad de las
posteriores, y estaban adaptadas para sujetar los alimentos.
El cuerpo era muy corto y la larga cola servía para mantener
el equilibrio. Leptictidium comía mucho más
que sólo insectos: algunos esqueletos presentan restos de
huesos de mamíferos pequeńos, huesos de lagartos y plantas,
además de fragmentos del esqueleto externo de algunos
insectos.
Europa (Francia). Tenia 1,5 m de longitud.
Potamotherium es la primera nutria de la que se
tienen noticias y, a semejanza de sus equivalentes modernos,
tenía el cuerpo largo y sinuoso y las patas cortas. Es
probable que corriera entre la vegetación baja de las
márgenes de los ríos, dando varios saltos, con la espalda
arqueada y la cabeza próxima al suelo. No tenía demasiado
desarrollado el sentido del olfato, pero parece que el oído
y la vista eran certeros y le servían para atrapar peces en
el agua. Sin duda, Potamotherium era un
excelente nadador; con su forma esbelta e hidrodinámica, no
debió de tener problemas para atravesar el agua, y su
columna vertebral flexible le permitiría zambullirse y
lanzarse con facilidad bajo la superficie. Las nutrias son
los únicos mustélidos que están bien representados en el
registro fósil. Esto se debe, con toda probabilidad, al
hecho de que vivían cerca del agua y, por lo tanto, era más
fácil que quedaran enterradas entre los sedimentos y después
se fosilizaran. Los paleontólogos piensan que las focas
auténticas, o fócidos, evolucionaron a partir de un
antepasado mustélido.
Egipto y America del Norte. No es el lugar
donde usted buscaría ballenas, pero en un tiempo el desierto
de Wadi Hitan estaba bajo agua e hirviendo con gigantes del
mar. En 2005 el geólogo Philip D. Gingerich anunció que su
equipo había extraído el primer esqueleto casi completo
conocido de un Basilosaurus isis. El fósil de
40 millones de ańos y de 50 pies de largo (18 metros de
largo) se enviará ahora a Michigan dónde los expertos lo
conservarán. Después ellos devolverán el fósil a Egipto
junto con un molde completo del esqueleto. La primera de las
ballenas verdaderamente gigantescas, Basilosaurus
tenía la forma de una serpiente o monstruo del mar con
dientes cortos y afilados para cazar tiburones y otras
presas. Al contrario de las ballenas de hoy, no tenía ningún
respiradero en el tope del cráneo — el antiguo monstruo
tenía que levantar la cabeza por sobre el agua para
respirar. Lo que es más, Basilosaurus todavía
tenía las patas y pies que heredó de sus antepasados que
moraban en la tierra, según Gingerich que trabaja para la
Universidad de Michigan con un subsidio de la National
Geographic Society. Cuando se encontraron por primera vez
los restos de esta sorprendente ballena primitiva, durante
la década de 1830, se creyó que correspondían a algún tipo
de dinosaurio. Esta criatura, que pertenece a la misma
familia que Zygorhiza, debió de tener el
aspecto de una enorme serpiente marina. Incluso se
utilizaron sus huesos en una broma famosa sobre una
serpiente de mar, hace alrededor de un siglo. El cuerpo
serpentino de Basilosaurus se apoyaba en una
columna formada por vértebras sumamente alargadas. Las
costillas eran cortas y estaban limitadas a la parte
anterior del cuerpo. Tambien se conoce a Basilosaurus
cetoides.
Asia (Pakistán). Tenia 1,8 m de longitud.
Pakicetus es la ballena más antigua que se conoce. Aunque
sólo se ha encontrado una parte de su cráneo, éste presenta
características tan primitivas que parece seguro suponer que
el resto de su cuerpo estaba poco adaptado para la
existencia marina.
Es probable que Pakicetus fuera muy diferente de las
ballenas modernas. Sus dientes eran similares a los de los
mesoníquidos, como Andrewsarckus, y las
muelas tenían la misma distribución triangular de las
cúspides. Esto sugiere que Pakicetus había evolucionado a
partir de los ungulados terrestres carnívoros apenas un poco
antes.
Sus orejas no estaban demasiado bien adaptadas para
funcionar debajo del agua, por lo que es probable que
Pakicetus pasara la mayor parte del tiempo en tierra. Los
descubrimientos de otros animales de vida terrestre en los
mismos depósitos en los que se ha hallado esta ballena
primitiva parecen confirmar esta teoría.
Se supone que Pakicetus tenía aspecto de foca. Las
extremidades habrán tenido forma de aletas, por lo cual sus
movimientos habrán sido torpes en tierra, pero lo hacían
sentir sumamente cómodo en los ríos y los estuarios a lo
largo de las costas orientales del Mar de Tetis. Esta
extensión de agua todavía existía a lo largo del extremo
meridional de Asia, al principio del Terciario, hace unos 50
millones de ańos.
Asia (Mongolia).
Tenia 3 m de longitud. Hace unos 35 millones de ańos,
durante el final del Eoceno, Asia Central contaba con
algunos mamíferos inmensos, como los bromoterios, los
calicoterios y los rinocerontes. Para poder aprovechar unas
presas tan impresionantes, los creodontos aumentaron también
de tamańo. Uno de los más inmensos fue Sarkastodon,
más grande que los osos más enormes. Los dientes eran vastos
y tan gruesos como los del oso gris actual. También como los
osos modernos, es probable que Sarkastodon
comiera una gran variedad de alimentos. Entre los demás
oxiénidos que vivían en el hemisferio norte durante el
Paleoceno y el Eoceno, hace entre 55 y 40 millones de ańos,
podemos incluir unos animales que se parecían a los glotones
y los felinos.
Indias Occidentales
(Jamaica). Los sirénidos, o vacas marinas, son el
único grupo de mamíferos que se han convertido en herbívoros
acuáticos completamente adaptados. En la actualidad, están
representados por tres especies de manatíes (Tri-chechus)
y una sola especie de dugongos (Dugong dugon).
Todos tienen cuerpos bulbosos, miembros anteriores
modificados en forma de aletas, carecen de extremidades
posteriores y tienen una cola aplanada en sentido
horizontal, como la dé la ballena, que utilizan para
impulsarse a través del agua a un ritmo mesurado. Los
sirénidos se conocen desde el comienzo del Eoceno en
Hungría. Su evolución es misteriosa, pero muchos
paleontólogos creen que es posible que compartan un
antepasado con los elefantes. Durante todo el Eoceno, el
clima fue bastante templado y en las aguas tropicales y poco
profundas del Mediterráneo y el Caribe abundaban los prados
vastos cubiertos de hierbas marinas, el principal alimento
de los sirénidos marinos. Posiblemente 1,5 m de longitud. Prorastomuses el sirénido más primitivo que se
conoce. Hasta ahora, sólo se han descubierto su cráneo y
algunas partes de la columna vertebral y las costillas: por
lo tanto, la restauración que figura aqui, es una
especulación. El aspecto de su cráneo indica que no estaba
especializado para llevar una vida acuática y es probable
que Prorastomus todavía fuese, en esencia, un
habitante de la tierra. Su hocico grueso y sus muelas de
doble cresta indican una alimentación a base de vegetales
blandos.
Asia
(Mongolia). Cuando los
mamíferos herbívoros (sin contar los multituberculados)
aparecieron por primera vez, al comienzo del Paleoceno, no
existía ningún carnívoro que pudiera apresarlos. A mediados
del Paleoceno, sin embargo, hace más de 60 millones de ańos,
una raza primitiva se había transformado en un nuevo orden,
los acreodos. Entre ellos estaban los mesoniquidos, es
decir, omnívoros con aspecto de lobos, hienas u osos,
capaces de sacar provecho de esta nueva fuente de alimentos.
Su tamańo iba desde el del zorro hasta el inmenso
Andrewsarchus. Los mesoniquidos prosperaron hasta el
comienzo del Oligoceno, hace unos 35 millones de ańos,
cuando los creodontos, y después los carnívoros auténticos,
ya se habían convertido en los carnívoros dominantes. Las
similitudes en la distribución de los huesos en la base del
cráneo y en los dientes indican que, a pesar de las enormes
diferencias de hábitats y formas de vida, los mesoniquidos
tal vez-dieron origen a las ballenas y los delfines. Tenia 4
m de longitud. Con un cráneo gigantesco, de casi 1 m de
largo, Andrewsarchus fue el mamífero carnívoro
terrestre más grande que se haya conocido. Los dientes eran
enormes y estaban adaptados para triturar y desgarrar los
alimentos. La forma de vida de Andrewsarchus
sigue siendo un misterio, ya que nunca se ha encontrado un
esqueleto completo. Si lo comparamos con sus familiares,
resulta que no era un cazador que persiguiese a sus presas,
sino un carrońero, como la mayoría de las hienas actuales.
Asia.
Es un género extinto de serpientes que incluye una única
especie: Gigantophis garstini, que significa
"serpiente gigante de Garstin". Gigantophis
vivió hace aproximadamente 40 millones de ańos en el sur del
actual Sahara, en donde ahora se sitúan Egipto y Argelia.
Como indica su nombre, ésta era una serpiente de tamańo muy
grande: la segunda serpiente más grande de todos los tiempos
entre las conocidas, sólo superada por Titanoboa
cerrejonensis. Jason Head, del Instituto
Smithsoniano en Washington, D.C., ha comparado las vértebras
fósiles de Gigantophis con las de las
serpientes modernas, concluyendo que esta serpiente extinta
podía crecer a una longitud de entre 9.3 metros a 10.7
metros de longitud. De ser así había sido hasta un 10 por
ciento más larga que las especies mayores de la actualidad,
la anaconda verde y que la pitón reticulada. A pesar de su
tamańo, no se sabe todavía cuál era su hábitat, si marino o
terrestre. Sin embargo, se ha sugerido que, al igual que la
anaconda, tal vez fuese una moradora de los pantanos y
buscase presas del tamańo de una gacela grande o del de una
vaca, como el proboscídeo primitivo Moeritherium.
Se ha discutido también si esta serpiente tenía miembros
posteriores (pero no espolones como los que poseen las boas
y las pitones).
Ameghino F,
1891, ‘Nuevos restos de mamíferos fósiles
descubiertos por Carlos Ameghino en el Eoceno
inferior de la Patagonia austral. Especies nuevas,
adiciones y correcciones’, Revista Argentina de
Historia Natural, 1: 289-328.
Bellosi, E.S. y Genise, J.F. 2004.
Insect trace
fossils from paleosols of the Sarmiento Formation
(Middle Eocene-Lower Miocene) at Gran Barranca (Chubut
Province). First International Congress on Ichnology
(Trelew), Fieldtrip Guidebook, 15-29 pp.
Chimento N.R,
Agnolin F.L y Novas F.E(ed.), 2012, ‘The
Patagonian fossil mammal Necrolestes: a Neogene
survivor of Dryolestoidea’, Revista del Museo
Argentino de Ciencias Naturales, nueva serie, 14, 2.
Cladera, G., Ruigomez,
E., Ortiz Jaureguizar, E., Bond, M., & López, G.
(2004). Tafonomía de la Gran Hondonada (Formación
Sarmiento, Edad-mamífero Mustersense, Eoceno Medio)
Chubut, Argentina. Ameghiniana, 41(3), 315-330.
Madden, R.,
Carlini, A., Vucetich, G., Kay, R., Heizler, R.,
Vilas, F., Ré, G., Kohn, M., Zucol, A. y Bellosi, E.
2003. Gran
Barranca: The most complete South America middle
Cenozoic sequence.
Symposium on Paleogene
(Leuven, 2003),
Abstracts and Program, pp.
Magnussen Saffer,
Mariano (2012).
Vertebrados fósiles del Periodo Eoceno de la Republica
Argentina. Paleo, Revista
Argentina de Paleontología. Boletín Paleontológico. Ańo 10.
76: 21-28.
Magnussen Saffer,
Mariano (2013). Mamíferos marinos
característicos del Eoceno. Paleo, Revista Argentina de Paleontología. Boletín
Paleontológico. Ańo 11. 91: 24-28.
Magnussen Saffer,
Mariano (2013). Algunos
vertebrados continentales representativos durante el Eoceno.
Paleo, Revista Argentina de Paleontología. Boletín
Paleontológico. Ańo 11. 92: 23-31.
Pascual,
R. y Odreman Rivas, O. 1971. Evolución de las
comunidades de los vertebrados del Terciario
argentino. Los aspectos paleozoogeográficos y
paleoclimáticos relacionados. Ameghiniana 8:
372-412.
Simpson, G. G.,
Minoprio, J. L. y Patterson, B. (1962).
The mammalian fauna of the
Divisadero Largo Formation, Mendoza, Argentina.
Bulletin of the Museum of Comparative Zoology at
Harvard College 127: 237-303.
Ver mas
bibliografía utilizada para hacer la presente Pagina;
AQUÍ.
Ver bibliografía de divulgación sugerida; AQUÍ.
Video; Fieras
de la Prehistoria - Documental - National
Geographic