Descripción, características y
paleoecolologia de los principales reptiles
voladores (Pterosauria, Pterodactyloidea) del
Conosur.
Por Mariano Magnussen Saffer. Director
de Grupo Paleo e Integrante del Museo Municipal
Punta Hermengo.
marianomagnussen@yahoo.com.ar. Tomado de: Magnussen Saffer,
Mariano (2012).
Descripción, características y paleoecolologia de los
principales reptiles voladores (Pterosauria,
Pterodactyloidea) del Conosur. Paleo, Revista
Argentina de Paleontología. Boletín Paleontológico. Ańo 10.
80: 17-29.
Los pterosaurios (Pterosauria, gr. "lagartos
alados") son un clado de saurópsidos arcosaurios
voladores que existieron durante casi toda la
Era Mesozoica (228 a 65 millones de ańos).
Fueron los primeros vertebrados en conquistar el
aire. Sus alas estaban formadas por una compleja
membrana sostenida por el cuarto dedo de la
mano, que estaba hipertrofiado. Su cuerpo estaba
cubierto de pelo, preservado en fósiles de al
menos tres especies distintas. Los fósiles de
pterosaurios son muy comunes y se han hallado
centenares de especímenes en África, Asia,
Australia, Europa, Norteamérica y Sudamérica.
Los pterosaurios no son dinosaurios, aunque sí
convivieron con ellos durante decenas de
millones de ańos. El término "dinosaurio" está
restringido a ciertos grupos de reptiles con una
postura erguida única (el superorden Dinosauria,
el cual incluye a las aves), y por lo tanto
excluye a los pterosaurios, así como a varios
grupos de reptiles marinos extintos como los
ictiosaurios, plesiosaurios y mosasaurios.
 |
Los pterosaurios son también
frecuentemente denominados como
"pterodáctilos", particularmente por
periodistas. Este uso es desaconsejado.
"Pterodáctilo" se refiere
específicamente a los miembros del
género Pterodactylus, y más
ampliamente a los miembros del suborden
Pterodactyloidea. Los pterodactiloideos,
son uno de los dos subórdenes de
pterosaurios o reptiles voladores.
Reciben el apelativo de pterosaurios de
cola corta, en contraposición a los
miembros del otro suborden, los
ranforrincoideos (pterosaurios de cola
larga). |
A veces reciben el nombre común de
pterodáctilos, término que en ocasiones se
aplica también a todos los pterosaurios, y que
otras veces se usa para referirse solo al género
Pterodactylus. Entre los ejemplos más
conocidos de pterodáctilos se encuentran
Pterodactylus, Dsungaripterus, Pteranodon y
Quetzalcoatlus.
Los pterodactiloideos se diferencian de los
ranforrincoideos, más basales (primitivos), por
la cola corta y los metacarpianos del ala
largos; los más derivados carecen de dientes.
Muchas especies tuvieron crestas bien
desarrolladas en el cráneo, que alcanzaron su
máximo tamańo en géneros como Nyctosaurus
y Tupandactylus.
Tropeognathus (=Ornithocheirus).
Wellnhofer, 1987.
Es un género de pterosaurios pterodactiloideos
de la familia Ornithocheiridae. Fue un
pterosaurio de grandes dimensiones que habitó
Sudamérica y Europa en el Cretácico Inferior.
Basado en restos fósiles escasos, el género ha
causado problemas duraderos de nomenclatura
zoológica.
Los restos fósiles actualmente clasificados como
Ornithocheirus han sido recuperados
mayormente de Cambridge Greensand en Inglaterra,
datando de principios del Albiano en el período
Cretácico, hace unos 110 millones de ańos.
Fósiles adicionales de la formación Santana de
Brasil, datando de entre 112 a 108 millones de
ańos, han sido clasificados como una especie de
Ornithocheirus.
 |
El material original de Ornithocheirus simus,
recuperado de Inglaterra, indica una especie de
tamańo medio con una envergadura de 2.5 metros.
Espécimenes referidos atribuidos a
Ornithocheirus simus (alternativamente
llamado Criorhynchus simus) pueden
alcanzar los 5 metros. Tropeognathus
mesembrinus es también usualmente
considerado como parte del género
Ornithocheirus como O. mesembrinus, y
alcanzaba cerca de 6 metros de envergadura. |
Tanto O. simus como O. mesembrinus
tenían distintivas crestas convexas en forma de
quilla sobre sus hocicos. Las crestas superiores
parten desde la punta del hocico y se extienden
hasta la fosa nasal. Una cresta menor adicional
se proyectaba hacia abajo desde la mandíbula
inferior en la sínfisis (el área del "mentón").
Aunque muchos ornitoqueíridos tenían una cresta
ósea redondeada proyectándose en sus hocicos,
esta era particularmente grande y bien
desarrollada en Ornithocheirus.
A diferencia de sus parientes Anhanguera
y Coloborhynchus, que tenían una roseta
de dientes en la punta de sus mandíbulas,
Ornithocheirus tenía mandíbulas rectas que
se estrechaban hacia la punta. A diferencia de
otros pterosaurios emparentados, los dientes de
Ornithocheirus eran casi verticales, en
vez de situarse en un ángulo abierto. También
tenía menos dientes que sus demás parientes.
 |
El espécimen tipo de Ornithocheirus simus
es representado sólo por una pieza rota de la
punta de la mandíbula superior. Aunque ésta
preserva varios rasgos característicos de
Ornithocheirus, es casi idéntica a los
huesos comparables en o. mesembrinus,
haciendo que una distinción clara entre estas
dos especies sea imposible.
Como resultado, aunque cerca de cuarenta
especies habían sido nombradas en el género
Ornithocheirus a través de los ańos, ni una
sola de ellas, ni siquiera O. simus, es
actualmente reconocida como válida por todos los
expertos en pterosaurios. |
Frecuentemente, hay
una carencia total de consenso; por ejemplo
Tropeognathus mesembrinus nombrado por Peter
Wellnhofer en 1987 ha sido posteriormente
considerado como Ornithocheirus mesembrinus
por David Unwin en 2003 (haciendo a
Tropeognathus un sinónimo menor), pero
también como Anhanguera mesembrinus por
Alexander Kellner en 1989, Coloborhynchus
mesembrinus por André Veldmeijer en 1998 y
Criorhynchus mesembrinus por Michael
Fastnacht en 2001. Incluso antes, en 2001, Unwin
había referido el material de "Tropeognathus" a
O. simus lo cual fue seguido por
Veldmeijer; sin embargo este último negó que
O. simus sea la especie tipo en favor de
O. compressirostris (alternativamente
Lonchodectes), y usó los nombres
Criorhynchus simus y Cr. mesembrinus.
Kellner en 2000 de nuevo reconoció a
Tropeognathus como un género válido.
Anhanguera
santanae.
(Wellnhofer, 1985
Es un género de pterosaurios pterodactiloideos
conocidos por la formación brasileńa de Santana
del Cretácico inferior (Aptiano). Debe su nombre
a la ciudad brasileńa de Anhangüera que a su vez
recibió el nombre de una deidad amerindia
protectora de la naturaleza. El descubrimiento
de este pterosaurio ayudó a finalizar el debate
sobre si los pterosaurios caminaban a dos o
cuatro patas.
 |
Los Anhanguera eran criaturas piscívoras,
como indican sus dientes, y buenas voladoras,
con una envergadura de alas de entre 4 y 5 m por
sólo uno y medio de alto, lo que los convertía
en unos de los mayores pterosaurios de la época.
Tenían una pequeńa cresta redondeada en la parte
frontal de sus mandíbulas que le daban un
aspecto característico.
Estos pterosaurios están cercanamente
emparentado con el ornitoqueiro, y
pertenecen a su misma familia Ornithocheiridae
en una subfamilia propia, Anhanguerinae, que
también incluye al Ludodactylus.
Hay varias especies de Anhanguera
reconocidas con una amplia distribución. A.
santanae y A. blittersdorfi se
conocen por varios fósiles fragmentarios que
incluyen esqueletos de la formación de Santana
de Brasil.
|
A. cuvieri y A. fittoni,
que inicialmente se habían descrito
pertenecientes al género Pterodactylus y
posteriormente a Ornithocheirus, son de
un periodo un poco posterior al (Albiano) en
Inglaterra, mientras que fragmentos de un
pterosaurios que puede atribuirse a
Anhanguera se han encontrado en Queensland,
Australia. La especie anteriormente conocida
como A. piscator ha sido incluida en un
género diferente, Coloborhynchus, (Veldmeijer,
2003).
Un estudio de 2003 se afirma que los
Anhanguera mantenían su cabeza inclinada
respecto a la horizontal, según muestra la
estructura de su oído interno, que ayudaba al
animal a mantener el equilibrio de forma similar
a como ocurre en los humanos.
Pteraichnus s.p.
Stokes, 1957.
Hallado en la Patagonia y en otras partes del
mundo, podría tener algún aspecto similar a los
Pterodaustro del Jurasico de San Luis. La única
evidencia que existe hasta hoy, son unas lajas
pertenecientes a sedimentos costeros en donde
quedaron impresas las huellas de este raro
animal, lo que no le permite a los Paleontólogos
hacer una reconstrucción fidedigna. Las huellas
se encuentran depositadas en el Museo de
Geología y Paleontología de la Universidad
Nacional del Comahue. Los restos fósiles de este
animal proceden del Cretácico de la Villa El
Chocon en la Provincia de Neuquen, Argentina. Su
antigüedad es de 105 millones de ańos.
Los
Pteraichnus
icnogénero
por ejemplo,
inicialmente
definidas
como impresiones
pterosaurio
pterodactyloid
desde
el Jurásico Tardío al
Cretácico Tardío.
 |
De hecho, este
intervalo de
tiempo es
poco probable que
refleje la
existencia de un
solo género, es
decir que este
icnogénero
incluye
impresiones realizadas
por diferentes
especies, géneros
e incluso familias.
Stokes
(1957) a
diagnosticado a
Pteraichnus
como un
reptil volador de
andar
cuadrúpedo en tierra firme.
Manos
con
3
dígitos,
2
de
los cuales son
pequeńos y
llevan garras
cortas y romas;
la
tercera,
mucho más grande,
soporta
la membrana del ala
y
es capaz de ser
doblada
hacia atrás
y hacia arriba
a fin de dejar
sólo
un
breve impresión
posterior. |
Los pies
tiene 4
dígitos.
El talón
es muy estrecha,
la
única forma de V y
los
dedos de los pies
delgadas con
garras
curvas.
La longitud
inferirse
del cuerpo
exclusivo
de
cuello, cabeza y cola,
aproximadamente 30
pulgadas.
Al caminar,
la
parte posterior del pie
sobrepasa la parte
delantera del pie que
indica la posibilidad
de la postura bípeda
ocasional.
La
aparente reducción de
dígitos en ambos
manos y pies
es distintivo y
es la razón principal
para la colocación del
animal en el
Pterodactyloidea.
En
1995,
Lockley et
al.
corrigió el diagnóstico
de la siguiente manera:
«Vía de la pista
ancha
de un animal cuadrúpedo
con
alargados, simétricos,
funcional-tetractil,
impresiones plantígrados
pies y
asimétricas
manos
tridáctilas.
Impresiones de manos
dígitos
IV,
alargada, curvada
y dirigido hacia atrás,
paralela al eje de
vía de la pista.
Manos
impresiones a menudo
más profundamente
impresionados
que las impresiones
pie.
Tapejara wellnhoferi.
Kellner,
1989
Es un género de pterosaurios pterodactiloideos de tamańo medio
con una enorme cresta en la cabeza que le
servía, posiblemente, para el cortejo. Vivió en
del Cretácico, hace 108 millones de ańos. Se han
encontrado esqueletos perfectamente conservados
en la Formación Santana, en Brasil.
Tapejara
vivía en la orilla de los lagos o en islas
interiores, y es probable que fueran voladores
lentos y no recorriesen grandes distancias. Se
alimentaban fundamentalmente de peces.
Comparaciones entre los anillos escleróticos de
Tapejara y de reptiles y aves modernos
sugieren que este debió de haber sido catemeral,
es decir activo a través del día durante cortos
intervalos.
 |
La especie tipo y única actualmente
reconocida como válida por la mayoría de
investigadores es T. wellnhoferi,
la cual es la más pequeńa de las
especies que han sido asignadas a
Tapejara y no preserva evidencia de
extensiones de tejido blando en la
cresta. El nombre de la especie es en
homenaje al paleontólogo alemán Peter
Wellnhofer. Una segunda especie,
denominada originalmente Tapejara
imperator (que significa
"emperador"), es mucho mayor y posee una
distintiva cresta compuesta de un largo
puntal, proyectándose desde la cresta
redondeada del hocico y la parte
posterior del cráneo, la cual apoyaba
una enorme cresta redondeada en forma de
vela hecha de queratina. |
Una tercera especie, Tapejara navigans
(que significa "navegante"), era de tamańo medio
y mostraba una cresta similar a la de T.
imperator, aunque más estrecha y en forma de
domo, que carecía del puntal óseo de apoyo
orientado hacia atrás.
 |
Varios estudios publicados en 2007 mostraron que
T. imperator y T. navigans eran
demasiado diferentes de T. wellnhoferi y
por lo tanto requerían sus propios nombres de
género. A la especie T. imperator se le
reclasificó en su propio género,
Tupandactylus, por Kellner y Campos. Unwin y
Martill encontraron que T. imperator y
T. navigans pertenecían al mismo género, y
los nombraron Ingridia imperator e I.
navigans, respectivamente. El nombre del
género era en honor de la difunta esposa de
Wellnhofer, Ingrid. Debido a que
Tupandactylus fue nombrado primero, este
retiene la prioridad sobre el nombre Ingridia.
|
Para complicar las cosas, tanto el nombre
Tupandactylus como Ingridia usaban al
antiguo Tapejara imperator como su
especie tipo. Los científicos que describieron a
Tupandactylus no nombraron a
Tupandactylus navigans (pero en vez de ello
sugirieron hacerlo sinónimo de Tupandactylus
imperator), y Tapejara navigans no
fue formalmente reclasificado como una especie
distinta de Tupandactylus hasta 2011.
Tupuxuara longicristatus. Kellner &
Campos, 1988
Es
un género de pterosaurios pterodactiloideos de la
superfamilia Azhdarchoidea que vivió a
principios del Cretácico (Valanginiano y
Hauteriviano) en lo que hoy es Brasil.
Tupuxara
medía unos 2,5 metros de largo y 5,4 m de
envergadura. Su principal rasgo distintivo era
una gran cresta en su cabeza, más redondeada en
las hembras y ausente en los individuos que no
habían alcanzado la madurez sexual. Esta cresta
iba desde el hocico hasta detrás del cráneo. La
cabeza, que medía casi 1 m, era enorme a
comparación con el cuerpo, era muy liviana,
gracias a los agujeros en el cráneo.
 |
Tupuxuara
es un miembro del grupo Azhdarchoidea. Kellner
lo asignó a la familia Tapejaridae dentro de
Azhdarchoidea. Sin embargo, de acuerdo a algunos
análisis, Tupuxuara es más cercano a los
Azhdarchidae (el grupo que incluye al gigantesco
Quetzalcoatlus de Norteamérica) que a
Tapejara y sus parientes.
Se ha sugerido que
Tupuxuara era un devorador de peces en
las costas de Suramérica. Otras hipótesis
incluyen la posibilidad de que consumiera
frutas. |
Un espécimen subadulto descrito por David
Martill y Darren Naish de la Universidad de
Portsmouth en 2006 aún no tenía una cresta
completamente desarrollada, lo que apoya la idea
de que la cresta era un indicador de madurez
sexual.
 |
Las comparaciones entre los anillos escleróticos
de Tupuxuara y aves y reptiles modernos
sugieren que pueden haber sido diurnos.
La especie T. longicristatus fue
descubierta en la Formación Santana en 1988 por
Alexander Kellner y Diógenes Campos. La especie
tipo es Tupuxuara longicristatus.
El
nombre del género se refiere a un espirítu
familiar de la mitología de los Tupí. El nombre
de la especie significa "cresta alaragda" en
latín. |
En 1994 una segunda especie fue nombrada por
Kellner: Tupuxuara leonardii. El nombre
de la especie es en homenaje a Giuseppe Leonardi.
El holotipo es MN 6592-V, un cráneo fragmentario
con una cresta más redondeada. Otros materiales
han sido referidos a T. leonardii. Los
mayore cráneos tienen una longitud de 130
centímetros, indicando uan envergadura de 5.5
metros.
Thalassodromeus sethi.
Kellner & Campos, 2002.
Es
un género extinto de pterosaurios pterodactiloideos de la
superfamilia Azhdarchoidea. Fue un gran
pterosaurio que se encontró en el nordeste de
Brasil. Vivió hace aproximadamente entre 122 y
109 millones de ańos. Compartió los cielos con
su pariente más grande Tapejara.
El género fue nombrado en 2002 por Alexander
Kellner y Diogenes de Almeida Campos. La especie
tipo es Thalassodromeus sethi. El nombre
del género se deriva del griego thalasse,
"mar" y dromaios, "corredor", en
referencia a su presunto estilo de vida similar
a un rayador. El nombre de la especie se refiera
al dios egipcio Seth debido a la similitud en la
forma de sus cabezas. En 2006 André Jacques
Veldmeijer sugirió que Kellner había confundido
a Seth con el dios Amón cuya corona tiene un
considerable parecido con la cresta de la cabeza
de Thalassodromeus.
 |
El género está representado por un cráneo
parcial dańado, hallado en la Formación Santana.
Thalassodromeus vivió a principios del
Cretácico Superior, hace más o menos 108
millones de ańos. Este compartía los cielos con
su pariente menor Tapejara. Es
particularmente notable por su inmensa cresta
craneal, comenzando en la punta del hocico y
finalizando por detrás del neurocráneo, lo cual
hace el setenta y cinco por ciento de la
superficie de su cráneo de 1.42 metros de largo.
Las mandíbulas acababan en punta y eran
desdentadas.
Poseía una envergadura de aproximadamente 4.5
metros. La función de esta cresta es
desconocida, pero puede haber sido usada para
exhibición sexual, renocimiento de individuos de
la especie o termorregulación. Un fragmento de
mandíbula inferior referido a Thalassodromeus,
DGM 1476-M, indica un ejemplar aún mayor con una
envergadura de 5.3 metros. |
Otro fragmento de
mandíbula, SAO 251093, fue considerado de manera
no oficial como una nueva especie, "Thalassodromeus
oberli" (refiriéndose a la colección Urbs Oberli),
por Veldmeijer, pero más tarde él cambió de
opinión y refirió el espécimen a T. sethi.
Kellner creyó que Thalassodromeus se
alimentaba de forma muy similar al actual
rayador; arrastrando su mandíbula inferior en el
agua mientras volaba. Sin embargo, la
investigación posterior mostró que la anatomía
de la mandíbula y el cuello sugerían que para
este y otros grandes pterosaurios no era
factible rozar el agua de esa manera debido al
arrastre: el gasto de energía podría haber sido
demasiado alto.
Kellner asignó a Thalassodromeus a la
familia Tapejaridae. Otros análisis sin embargo,
han indicado que este estaba relacionado con
Tupuxuara en el grupo Thalassodrominae, más
cercanamente relacionado a la familia
Azhdarchidae.
Pterodaustro guinazui.
Bonaparte, Guińazú, 1979
Es
un género de pterosaurios pterodactiloideos del Cretácico
Superior que vivió en lo que hoy es Sudamérica
hace 140 m. a.
Pterodaustro
tenía una envergadura de 132 cm; su cabeza medía
23 cm de longitud y era muy estrecha con un pico
curvado en el extremo.
 |
Tenía una boca armada con más de 500
largos y angostos dientes, que
presumiblemente estaban diseńados para
filtrar el contenido alimenticio
suspendido en agua, de manera similar a
como lo hacen los modernos flamencos.
Poseía infinidad de diminutos rizos en
las mandíbulas que podrían haber sido
empleados para capturar crustáceos y
otras pequeńas criaturas acuáticas.
La dieta del animal podría haberle
conferido una tonalidad rosada, otro
probable rasgo en común con los
flamencos de hoy en día; en efecto,
Pterodaustro es frecuentemente
apodado el "pterosaurio flamenco". |
Fue descubierto en 1970 por los paleontólogos
José Bonaparte y José Román Guińazú en un paraje
perteneciente a la Formación Lagarcito, situada
en la provincia de San Luis, en el centro de
Argentina. Kevin Padian, paleontólogo de la
Universidad de California, junto a Michael Bell,
hallaron un yacimiento con un gran numero de
fósiles de este reptil volador en el norte de
Chile. Se sugiere que esta formación se originó
cuando sucesivas y repentinas inundaciones
arrasaron con nidales de este reptil. Fue el
primer pterosaurio hallado en el hemisferio sur.
Domeykodactylus ceciliae. Martill et al., 2000
Es
un pterodáctilo dsungariptérido del Cretácico
inferior que vivió en Chile. El género fue
descrito en el ańo 2000 por David Martill,
Eberhard Frey, Guillermo Chong Diaz y Charles
Michael Bell. La especie es Domeykodactylus
ceciliae. El nombre se deriva de la
Cordillera Domeyko y del griego daktylos,
"dedo", en referencia al alargado dedo en el
ala típico de los pterosaurios. El nombre de la
especie honra a la geóloga Cecilia Demargasso de
la Universidad Católica del Norte y su
colaboración con los autores.
 |
La descripción del Domeykodactylus se
basa en el ejemplar conservado en la Universidad
Católica del Norte de Antofagasta, encontrado en
la Sierra da Candeleros, Chile. Consiste de una
mandíbula parcial; un premaxilar, presente en la
misma roca, es referido como el paratipo. El
fósil fue en principio considerado como
perteneciente a Pterodaustro.
Domeykodactylus tenía una cresta que corría
a lo largo de la parte superior del premaxilar.
La estructura del hueso de la cresta consiste de
trabéculos verticales, que eran puntales
estrechos; es esta textura la que originalmente
fue confundida con los delicados dientres
filtradores de Pterodaustro.
La mandíbula tenía una sínfisis corta.
Tenía dieciséis alvéolos para los
dientes, de los cuales han desaparecido
los propios dientes, en ambos huesos
dentarios. Los alvéolos son estrechos,
ovales y levemente elevados, sobre el
nivel de la mandíbula. |
Los dientes eran
probablemente pequeńos y se hacían más pequeńos
y más espaciados entre sí hacia la parte posteriro de la boca. La longitud del cráneo se
ha estimado en treinta centímetros y la
envergadura alar en un metro.
Los descriptores encontraron que
Domeykodactylus era similar tanto a los
Ctenochasmatidae como a los Dsungaripteridae en
la cresta; debido a la posesión de alvéolos
dentales elevados fue asignado a este último
grupo. Fue el primer ejemplo publicado de un
dsungaptérido en América del Sur, mientras que
la mayoría de los otros miembros de la familia
han sido hallados en Asia
Cearadactylus
atrox. Leonardi & Borgomanero,
1985
es un género extinto de un pterosaurio grande
del Cretácico Inferior (Albiano) de Suramérica.
La especie tipo es Cearadactylus atrox,
descrita y nombrada en 1985 por Giuseppe
Leonardi y Guido Borgomanero. El nombre del
género se refiere al estado brasileńo de Ceará
combinándolo con el término griego daktylos,
"dedo", una referencia al dedo del ala de los
pterosaurios. El nombre de la especie significa
"feroz" en latín, una referencia a su temible
dentición.
El holotipo fue hallado en el Miembro Romualdo
de la formación Santana. Este fósil, un único
cráneo con una longitud de 57 centímetros, fue
descubierto en la meseta de Araripe en el
noreste de Brasil. Fue vendido a Italia en 1983
y adquirido por Borgomanero para su colección.
El cráneo está bastante dańado, especialmente en
la parte superior, y fue quizás reconstruido por
el vendedor de fósiles.
Como se mostró en una preparación realizada por
el Museu Nacional de Brasil, en la primera
preparación se cometieron varios errores graves.
El frente del hocico en ambas mandíbulas fueron
confundidos llevndo a una recosntrucción en la
que la parte anterior de la cabeza fue puesta al
revés. Los dientes fueron bastante restaurados y
alargados hasta el amplio frente de las
mandíbulas mostrándolos como muy grandes y
robustos y proyectándose hacia afuera, formando
una especie de "roseta".
 |
Esta mandíbula superior retorcida y sus dientes
entrelazados sugieren una dieta piscívora,
permitiéndole al animal mantener la sujeción de
peces resbaladizos. No parecía que hubiera
alguna cresta. La nueva prepraración hizo claro
que una cresta estaba presente sobre el hocico y
que la roseta era mucho menor. Muchos detalles
que fueron descubiertos resultaron útiles para
determinar la posición filogenética de
Cearadactylus.
La envergadura de Cereadactylus fue
estimada por los descriptores en cerca de 4
metros, con un peso de quizás 15 kilogramos.
Peter Wellnhofer en 1991 estimó una envergadura
de 5.5 metros.
|
Leonardi se abstuvo de asignarle una familia a
este género. Wellnhofer en cambio creó una
familia monotípica, Cearadactylidae, pero este
concepto ya no se usa. En 2000 Alexander Kellner
concluyó que estaba emparentado con los
Anhangueridae sin formar parte de ellos, ya que
carecía de cresta, dentro de un Pteranodontoidea
más amplio sensu Kellner. En 2002 David
Unwin sin embargo que era un miembro muy
modificado de Ctenochasmatidae. En 2010 Kellner
ingresó la nueva información en tres bases de
datos existentes de rasgos de pterosaurios, para
calcular a través de la cladística la posición
de Cereadactylus en un árbol
filogenético. Aunque los tres árboles diferían,
todos tenían en común la cercanía de
Cereadactylus a los Anhangueridae.
Cearadactylus ligabuei.
Dalla Vecchia 1993.
En 1993 Fabio Marco Dalla Vecchia nombró una
segunda especie, Cearadactylus ligabuei.
El nombre de la especie es un homenaje a
Giancarlo Ligabue, el director del Centro Sudi
Ricerche Ligabue en Venecia. Está basada en el
holotipo CCSRL 12692/12713, de nuevo un cráneo
sin cresta muy dańado, de 403 milímetros de
largo.
 |
El cráneo consiste de dos piezas, la
frontal y la posterior, unidas por los
vendedores de fósiles; se desconoce si
estos pertenecían a un solo individuo o
incluive si a una misma especie. El
propio Dalla Vecchia no estaba
convencido de que la nueva especie en
realidad perteneciera a Cearadactylus,
pero el cráneo no tenía suficientes
diferencias como para formar un nuevo
género y aún así era demasiado diferente
de las especies conocida como para
asignarlo a éstas, entonces él creó una
nueva especie para el género al que el
fósil más se parecía. Autores
posteriores ha negado la identidad
referida de este taxón denominándolo
como "Cearadactylus" ligabuei. |
Dalla Vecchia estimó la envergadura en seis metros;
Kellner, seńalando que le cráneo no era amyor que el
del holotipo de C. atrox', la estableció en cinco
metros como mucho. Dalla Vecchia asignó a C.
ligabuei a los Cearadactylidae. Kellner concluyó
que era probablemente un miembro de Anhangueridae;
Unwin en 2002 incluso lo denominó Anhanguera
ligabuei. Steel et.al. (2005) sugirió que debía
ser clasificado como Coloborhynchus ligabuei.
Nyctosaurus gracilis (=Pteranodon). Marsh, 1876.
Nyctosaurus es un género extinto de pterosaurio pterodactiloide de la
familia de los nictosáuridos, cuyos restos han sido
hallados en la Formación Niobrara en el medio oeste
de Estados Unidos, el cual, durante el período
Cretácico Superior, estaba cubierto por un extenso
mar poco profundo. El género Nyctosaurus ha
tenido numerosas especies clasificadas en este,
aunque cuantas de estas son realmente válidas es
algo que requiere de investigación adicional. Al
menos una de las especies poseía una extraordinaria
cresta craneal de gran tamańo y en forma de asta.
 |
Nyctosaurus
era similar anatómicamente a su pariente cercano
contemporáneo, Pteranodon. Ambos tenían alas
relativamente alargadas, similares en forma a las de
las aves modernas. Sin embargo, era mucho menor que
Pteranodon, alcanzando una envergadura en la
adultez de 2 metros y un peso máximo de cerca de
1.86 kilogramos. La longitud total del cuerpo era de
37 centímetros. Algunos especímenes preservan una
cresta distintiva, de al menost 55 centímetros de
alto en los especímenes de mayor edad, relativamente
gigante comparada con el resto del cuerpo y
cerca de tres veces mayor que la longitud de
la cabeza. |
La
cresta se compone de dos largas varas estriadas, una
apuntando hacia arriba y otra hacia atrás, partiendo
de una base común que se proyecta hacia arriba y
hacia atrás de la parte posterior del cráneo. Ambas
varas eran casi iguales en longitud, y ambas eran
tan largas o más que la longitud total del cuerpo.
La vara que apuntaba hacia arriba era de al menos
42 centímetros de largo y la que apunta hacia atrás
mide al menos 32 centímetros de largo.
Las
mandíbulas de Nyctosaurus eran largas y
extremadamente puntiagudas. Las puntas de las
mandíbulas eran delgadas y aguzadas como un alfiler,
y frecuentemente se encuentran rotas en los
especímenes fósiles, dando la apariencia de que una
mandíbula es más larga que la otra, aunque en vida
era probablemente iguales en longitud.
Nyctosaurus
es el único pterosaurio conocido que perdió sus
"dedos" con garras, con la excepción del dedo alar
(del cual sin embargo tampoco permanecía la cuarta
falange), lo cual probablemente habría afectado su
capacidad de desplazarse sobre tierra, llevando a
los científicos a conjeturar que pasaba casi todo si
tiempo en el aire y raramente aterrizaba. La
carencia de garras con las cuales poder aferrarse a
superficies pudo haber hecho particualrmente
imposible que trepara a los árboles o se sujetara a
acantilados.
Todos los fósiles conocidos de Nyctosaurus
provienen de la Caliza Smokey Hills en Kansas, parte
de la Formación Niobrara.
 |
Más exactamente, han sido
hallados solo dentro de una estrecha zona
caracterizada por la abundancia de fósiles de ammonites perteneciente a la especie Spinaptychus
sternbergi. Estos depósitos de calizas fueron
depositados durante una regresión marina del Mar
Niobraran que se produjo entre 85 y 84.5 millones de
ańos. Por lo tanto, Nyctosaurus fue un género
de escasa duración, a diferencia de su pariente
Pteranodon, el cual es encontrado a través de
casi todos los estratos de Niobrara hasta la
sobrepuesta Formación Pierre Shale, y existió entre
hace 88 a 80.5 millones de ańos. |
El
ecosistema preservado en esta zona era único por su
abundancia de vertebrados presentes. Nyctosaurus
compartía los cielos con el ave Ichthyornis y
con Pteranodon longiceps, aunque la otra
especie de Niobrara de Pteranodon, P.
sternbergi (denominada también Geosternbergia),
había desaparecido ya del registro fósil por esa
época. En las aguas del Mar Niobraran nadaban
mosasáuridos (como Clidastes, Ectenosaurus
y Tylosaurus), el ave no voladora buceadora
Parahesperornis, y una amplia variedad de
peces, incluyendo al Protosphyraena similar a
un pez espada, el depredador Xiphactinus y el
tiburón Cretolamna[4
Coloborhynchus (=Pterodactylus).
Owen, 1874.
Coloborhynchus es un género de pterosaurios pterodactiloideos de la
familia Ornithocheiridae del Cretácico Inferior en
Europa, América del Norte y América del Sur (época
del Albiano, hace 98 millones de ańos), y
posiblemente el Aptiano (hace entre 112 a 99
millones de ańos) de Brasil y Texas, dependiendo de
que especies sean incluidas. Posee varias especies,
algunas asignadas en un principio a otros géneros,
todas basadas en cráneos o en restos parciales.
El
espécimen tipo de Coloborhynchus es conocido
sólo a partir de una mandíbula superior parcial. Por
lo tanto, de acuerdo a la revaluación de Rodrigues y
Kellner de 2008 sobre Coloborhynchus clavirostris,
este solo puede ser diferenciado de sus parientes
basándose en su combinación única de posiciones de
los alvéolos dentales. En Coloborhynchus, los
dos dientes frontales apuntan hacia adelante y son
más altos sobre la mandíbula que los otros dientes,
mientras que los siguientes tres pares de dientes
apuntan a los lados. Los dos pares finales de
dientes (preservados) apuntan hacia abajo.
Finalmente, una única depresión oval estaba
localizada bajo el primer par de dientes.
 |
Como su pariente Anhanguera y
Uktenadactylus, la punta del hocico se ensancha
en una amplia roseta, en contraste con la estrecha
parte posterior de la mandíbula. También igual que
sus parientes, Coloborhynchus tenía una
cresta en forma de quilla en el frente de sus
mandíbulas, aunque esta era amplia y se adelgazaba
desde la base a a la cima, en vez de ser uniformente
delgada como en sus familiares. Esta clase de cresta
engrosada es también vista en Siroccopteryx
moroccensis, el cual puede ser su pariente más
cercano, si es que no era un miembro del mismo
género. También tenía un margen frontal recto, en
vez de ser curvado, a diferencia de otros de sus
parientes, y comenzaba en la punta del hocico, más
que de la parte posterior como en otras especies.
|
Un
segundo espécimen que muestra todas estas
características únicas fue reportado al paleontólogo
brasileńo Alexander Kellner por Darren Naish en
2007, y probablemente representa un segundo
espécimen de C. clavirostris, aunque aún no
ha sido descrito.
La
posible especie Coloborhynchus capito
representa el mayor ornitoqueírido conocido, y de
hecho es el mayor pterosaurio dentado que se
conozca. Un espécimen referido de Cambridge
Greensand en Inglaterra descrito en 2011 consiste de
una punta mandíbula superior muy grande que exhibe
las características dentales que distinguen a C.
capito de otras especies. La punta de la
mandíbula mide cerca de 10 centímetros de alto y 5.6
centímetros de ancho, con dientes de más de 1.3
centímetros de diamétro en su base. Si las
proporciones de este espécimen eran consistentes con
las de las otras especies conocidas de
Coloborhycnhus, la longitud total craneal pudo
haber sido de más de 75 centímetros, lo que implica
una envergadura estimada de 7 metros.
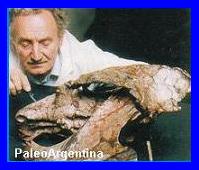
Aerotitán Sudamericanus.
Novas et al, 2012.
Poco antes de dar fin al presente articulo, recibi
la información del hallazgo de un nuevo reptil
volador, procedente de la Patagonia Argentina. El
pterosaurio que habitó Río Negro hace 65 millones de
ańos fue presentado en sociedad en el Museo
Patagónico de Ciencias Naturales. Los restos
constituyen el primer hallazgo en Sudamérica de un
reptil volador del Cretácico, la última de las eras
de los dinosaurios.
 |
El pterosaurio ha sido apodado como
Aerotitán Sudamericanus según su nombre en
latín, "titán del aire Sudamericano.
El animal sobrevoló los cielos de una
Patagonia bastante alejada de lo que hoy
conocemos. El ambiente era cálido y estaba
cubierto por un mar poco profundo que se
extendía hasta las proximidades de la
cordillera de los Andes. El continente
sudamericano ya se había separado de África,
había un Océano Atlántico más estrecho y la
península antártica estaba mucho mas cerca.
Tenía 10 metros de envergadura y era
carnívoro. Fue descubierto en diciembre 2010
durante una campańa argentino-sueca |
El
trabajo científico acaba de publicarse en la revista
norteamericana Journal of Vertebrate Paleontology,
siendo sus autores: Fernando E. Novas, Martin
Kundrat, Federico Agnolin, Martin Ezcurra, Per
Ahlberg, Marcelo Isasi, Alberto Arriagada y Pablo
Chafrat
De
acuerdo con la investigación, publicada en el
Journal of
Vertebrate Paleontology, se trataría del
único pterosaurio de la familia Azhdarchidae que fue
descubierto en la región. Los azhdárquidos incluyen
algunos de los reptiles alados más grandes que se
encontraron al momento, y dominaron los cielos hasta
la extinción de los dinosaurios.
“Este hallazgo es único en su tipo porque son los
primeros restos de pterosaurios de todo el
continente sudamericano correspondientes a fines de
la Era Mesozoica”, explica Fernando Novas,
investigador independiente del CONICET en el Museo
Argentino de Ciencias naturales (MACN-CONICET) y uno
de los autores del trabajo.
 |
En
diciembre de 2010 un grupo de paleontólogos de
Argentina y Suecia desenterraron los fósiles del
Aerotitan
en un yacimiento de la provincia de Río
Negro. A
pesar de que sólo encontraron un pico y fragmentos
de huesos de las alas, por sus características
pudieron determinar que se trataba de un ejemplar de
6 metros de largo de ala a ala y con un hocico largo
y delgado, similar al de una cigüeńa. |
“El
interior de los huesos preservan trabéculas óseas
delgadas, típicas de los pterosaurios”, analiza
Novas, “que les conferían fortaleza estructural pero
les permitían ser lo suficientemente livianos para
suspender sus cuerpos en el aire con unos pocos
aleteos”.
Según el paleontólogo, esto permite demostrar que
‘enormes pterosaurios’ sobrevolaron los mares
tropicales de la Patagonia a finales del Mesozoico.
“Este descubrimiento nos alienta a proseguir
explorando esta región de Río Negro, que podría
aportar datos de enorme interés para develar el
porqué de la desaparición de los grandes reptiles
hace 65 millones de ańos”.
Bibliografía Sugerida.
BILLON-BRUYAT, J-P.
& MAZIN, J-M. The systematic problem of tetrapod
ichnotaxa: the case study
of Pteraichnus Stokes, 1957 (Pterosauria,
Pterodactyloidea) página 319. GEOLOGICAL SOCIETY
SPECIAL PUBLICATION NO. 217 Evolution and
Palaeobiology of Pterosaurs.
Campos, D. A., and Kellner, A. W. A. (1985).
"Panorama of the Flying Reptiles Study in Brazil and
South America (Pterosauria/ Pterodactyloidea/
Anhangueridae)."
Anais da Academia Brasileira de Cięncias,
57(4):141–142 & 453-466
Campos, D. de A., and Kellner, A. W. (1985). "Un
novo exemplar de Anhanguera blittersdorffi (Reptilia,
Pterosauria) da formaçao Santana, Cretaceo Inferior
do Nordeste do Brasil." In Congresso Brasileiro de
Paleontologia, Rio de Janeiro, Resumos, p. 13.
Malam, J. & Parker, S. 2004. Enciclopedia de los
Dinosauros y otras criaturas prehistóricas.
Parragon Books Ltd, Bath.
Vullo, R. and Neraudeau, D. (2009). "Pterosaur
Remains from the Cenomanian (Late Cretaceous)
Paralic Deposits of Charentes, Western France."
Journal of Vertebrate Paleontology, 29(1):277-282.
Wellnhofer, P. (1991). The Illustrated Encyclopedia
of Pterosaurs. New York: Barnes and Noble Books. pp.
124.
Unwin, D. M. and Martill, D. M. (2007). "Pterosaurs
of the Crato Formation." In Martill, D. M., Bechly,
G. and Loveridge, R. F. (eds), The Crato Fossil Beds
of Brazil: Window into an Ancient World. Cambridge
University Press (Cambridge), pp. 475-524.
Unwin, D. M., (2003). "On the phylogeny and
evolutionary history of pterosaurs." In Buffetaut,
E. & Mazin, J.-M., eds. Evolution and Palaeobiology
of Pterosaurs.
London: Geological Society of London, Special
Publications 217, 2003, pp. 139-190.
Nuevas lecturas sobre el origen de
los mamíferos desde el Periodo Permico al Periodo
Cretácico.
Por Mariano Magnussen Saffer,
Director del Grupo Paleo. Presidente de la
Asociación Amigos del Museo Punta Hermengo.
marianomagnussen@yahoo.com.ar.
Tomado de;Magnussen Saffer,
Mariano (2010). Nuevas lecturas al origen de los mamíferos desde el Periodo
Permico al Periodo Cretácico. Paleo, Boletín
Paleontológico. Ańo 8. 44: 29-36.
Es
muy difícil hacer una rápida y breve mirada al
origen de los mamíferos, por lo cual, intentaremos
explicar lo mas significativo de este complejo
proceso. Parece unánimemente
aceptada la teoría de que los mamíferos proceden de
los reptiles, que evolucionaron para aprovechar
nichos ecológicos a los que antes no les era posible
adaptarse. La evolución de éstos a partir de los
sinápsidos (mamíferos como reptiles) fue un proceso
gradual que tuvo una duración de aproximadamente 70
millones de ańos entre el Pérmico Medio y el
Jurásico Medio, aconteciendo una gran explosión de
especies durante el Triásico Medio.
Sería correcto decir que
los mamíferos constituyen un grupo de animales con
columna vertebral, cuerpo aislado por pelo,
articulación única en la mandíbula, y que alimentan
a sus crías con leche. Sin embargo, esto no aclara
cómo estas pocas características comunes apoyan la
evolución de un grupo que presenta adaptaciones,
conductas, y sociedades muy complejas.
Los mamíferos son también
el grupo al que pertenecen los humanos, y a través
de ellos es mucho lo que podemos comprender acerca
de nosotros mismos. Por consiguiente otra respuesta
a la pregunta qué es un mamífero sería la de que su
esencia radica en su compleja diversidad de forma y
función, y sobre todo en su flexibilidad individual
de conducta, ya que el mamífero más pequeńo, el
murciélago de Kitti, pesa 1,5 g, en tanto que la
ballena azul pesa 100 millones de veces más; el lobo
puede viajar a travez de 1000 km2, y la
rata topo jamás abandona su madriguera; el opossum
de Virginia pare camadas de hasta 27 crías, y el
orangután sólo tiene un hijo; el elefante al igual
que el hombre, puede vivir unos setenta ańos,
mientras que el Antechinus stuari
macho jamás ve un segundo ańo, y muere antes de
nacer la primera y única camada de la que es
progenitor.
 |
Ninguna faceta de estas
vidas tan variadas sigue el azar: cada
mamífero potencia sus propiedades y
capacidades, a fin de dejar una descendencia
viable.
Los mamíferos son una
clase que comprenden unas 4.070 especies,
cuyas antiguas relaciones entre sí permiten
subdiviciones en unos 1.000 géneros, 135
familias, 18 órdenes y 2 subclases. Estas
últimas reconocen una separación de 200
millones de ańos entre los Prototheria
ovíparos (los únicos sobrevivientes son los
ornitorrincos y los equidnas) y los Theria
vivíparos. |
En cambio, solo tiene 90
millones de ańos la división entre los Theria, que
separa a los marsupiales (infraclase Metatheria) de
los mamíferos placentarios (Eutheria). Incluso
dentro de los compartimientos taxonómicos hay una
portentosa variación de tamańo, forma y modo de
vida: el comportamiento entre individuos de la misma
especie puede variar en razón de las circunstancias.
Así los individuos de una población de hienas
manchadas pueden formar clanes estables de unos 50
miembros o relacionarse sólo durante unos pocos
días.
Los mamíferos (Mammalia)
son una clase de vertebrados amniotas homeotermos
(de "sangre caliente"), con pelo y glándulas
mamarias productoras de leche con la que alimentan a
las crías. La mayoría son vivíparos (con la notable
excepción de los monotremas: ornitorrinco y
equidnas). Se trata de un taxón monofilético; es
decir, todos descienden de un antepasado común que
se remonta a finales del Triásico, hace más de 200
millones de ańos. Pertenecen al clado sinápsidos,
que incluye también numerosos "reptiles"
emparentados con los mamíferos, como los
pelicosaurios y los cinodontos
Los tetrápodos
Son animales vertebrados
provistos de cuatro extremidades o en su defecto
apéndices que se aproximan a ellas. En la actualidad
sólo existen cuatro grupos con especies vivas:
anfibios, reptiles, aves y mamíferos. También los
dinosaurios fueron tetrápodos. Una de las estirpes
de tetrápodos, evolucionó dando lugar a la aparición
de los amniotas. Se trata de los reptiliomorfos.
Se establecen como características de este grupo
animal un premaxilar estrecho, el vómer apuntando
hacia delante y una fórmula falangial (número de
uniones de cada dedo) en el pie.
Los Sinapsidos.
Reciben esta denominación
los tetrápodos que poseen una sola
cavidad craneal por encima de la línea de los huesos
temporales. A este grupo pertenecen además de los
fósiles que se detallan más abajo, todos los
mamíferos, incluido el hombre. Este grupo fue el
dominante de las faunas del período Pérmico, durante
el Triásico Inferior continuaron como grupo
dominante, pero a partir de la aparición de los
primeros
arcosaurios se produjo su declinación en número y
diversidad a partir del Triásico Medio, hasta su
casi extinción a fines del Triásico Superior. En
Ischigualasto están representados dos grupos de
sinápsidos, los cinodontes y los dicinodontes.
Los cinodontes.
Son un grupo de reptiles
mamiferoides cercanamente emparentados al origen de
los mamíferos, y uno de los principales componentes
de la paleofauna de Ischigualasto. Los cambios
evolutivos encaminados al estadío mamífero, se
reflejan en este tipo de animales en carácteres del
cráneo especialmente. Se caracterizaban por poseer
una dentición diferenciada con dientes incisivos,
caninos y molares. Además desarrollaron un extenso
paladar secundario, el que les permitía masticar
mientras respiraban, condición básica para mantener
un alto metabolismo. La mandíbula estaba formada
casi exclusivamente por el hueso dentario, ya que
los huesos posteriores que articulan con el cráneo
se habían reducido de tamańo. En los mamíferos estos
huesos han cambiado su función articular, a una
función auditiva, constituyendo los huesos del oído
medio. Los cinodontes que se han hallado en
Ischigualasto incluyen formas herbívoras y
carnívoras, y tamańos que varían desde el de un
ratón al de un jabalí grande.
 |
Los mamíferos actuales
descienden de los sinápsidos primitivos,
grupo de tetrápodos amniotas que comenzó a
florecer a principios del Pérmico, hace unos
280 millones de ańos, y continuaron
dominando sobre los «reptiles» terrestres
hasta hace unos 245 millones de ańos
(principios del Triásico), cuando empezaron
a despuntar los primeros dinosaurios. Debido
a su superioridad competitiva, estos últimos
hicieron desaparecer a la mayoría de los
sinápsidos. No obstante, algunos
sobrevivieron y se convirtieron en los
primeros mamíferos verdaderos hacia finales
del Triásico, hace unos 200 millones de
ańos. |
Los mamíferos más antiguos
que se conocen son, por un lado los
multituberculados y por otro los australosfénidos,
grupos que datan del Jurásico Medio. No obstante,
debe tenerse en cuenta que la organización mamaliana,
después de un éxito inicial durante el Pérmico y el
Triásico, fue suplantada casi por completo, en el
Jurásico y el Cretácico (durante unos 100 millones
de ańos), por los reptiles diápsidos (dinosaurios,
pterosaurios, cocodrilos, plesiosaurios,
ictiosaurios), y no fue hasta su desaparición cuando
los mamíferos se diversificaron y alcanzaron su
papel dominante. Aprovechar los recursos sin tener
que competir con animales de mayor envergadura
suponía adaptarse a regiones inhóspitas de clima
normalmente frío, a los hábitos nocturnos, también
con bajas temperaturas y además escasa iluminación.
A lo largo de la historia
evolutiva de los mamíferos acontecen una serie de
hechos que van a determinar la adquisición de los
rasgos que caracteriza a la clase. La capacidad
homeotérmica, es decir, de regular su temperatura
corporal, es sin duda alguna la característica que
permite a los mamíferos un mundo libre de
competencia y rico en recursos altamente nutritivos.
Fue gracias a ella que pudieron conquistar
territorios fríos y sobre todo, desarrollar una
actividad nocturna.
El crecimiento de pelo
protegiéndoles el cuerpo de la pérdida de calor y el
desarrollo de una visión apta para bajos índices de
luminosidad fueron las otras dos circunstancias que
colaboraron en la conquista de estos nichos
ecológicos hasta el momento libres de animales
superiores. Las adaptaciones del esqueleto fueron el
primer paso para conseguir mayor efectividad
energética basada en el incremento del
aprovechamiento de los recursos y en la disminución
del gasto. El cráneo va haciéndose más efectivo,
pierde masa, mantiene resistencia y simplifica
estructuras a la vez que permite el desarrollo y
efectividad muscular.
 |
Las modificaciones del
cráneo llevan además consigo la formación de
un paladar secundario, la formación de la
cadena ósea del oído medio y la
especialización de las piezas dentales.
La mandíbula se
constituye a partir de un único hueso (el
dentario) y ésta es la principal
característica para determinar si el fósil
de un animal pertenece a la clase de los
mamíferos, debido a la usual pérdida de
tejidos blandos durante la fosilización. |
Las extremidades dejan
paulatinamente de articularse a ambos lados del
tronco para hacerlo por debajo. De este modo, a la
vez que aumenta la movilidad del animal, disminuye
el gasto energético al hacer los requerimientos para
el desplazamiento y el mantenimiento del cuerpo
erguido. Por su parte, la gestación interna de las
crías y el proporcionarles a éstas los alimentos
para la primera edad sin tener que buscarlos
(leche), permitió mayor libertad de movimiento a las
madres y con ello un avance en su capacidad de
supervivencia tanto individual como de la especie.
En todos estos cambios
evolutivos se vieron involucradas todas y cada una
de las estructuras orgánicas, así como los procesos
fisiológicos. La maquinaria biológica
especializándose requería mayor efectividad de los
procesos respiratorios y digestivos, provocando el
perfeccionamiento de los aparatos circulatorio y
respiratorio con relación a la efectividad
fisiológica, y el del digestivo para conseguir un
mayor aprovechamiento nutritivo de los alimentos
fueron otros de los logros conseguidos por estos
animales durante su evolución.
El sistema nervioso central
fue adquiriendo un tamańo y estructura histológica
que no se conoce en otros animales, y la deficiencia
de iluminación a que se enfrentaban las especies
nocturnas se vio compensada con el desarrollo de los
otros órganos sensoriales, en especial del oído y el
olfato. Todos estos fenómenos evolutivos tardaron
varios cientos de millones de ańos, tras los cuales
los mamíferos hemos llegado a dominar la vida sobre
La Tierra, siendo las especies vivas con mayor grado
de evolución y desarrollo.
 |
La taxonomía clásica se
ha basado fundamentalmente en datos
morfológicos para establecer similitudes y
diferencias que permitan clasificar a las
distintas especies, pero los nuevos
descubrimientos paleontológicos y los
continuos avances en genética y biología
molecular ponen en entredicho bastantes de
las teorías evolutivas hasta el momento
aceptadas. Como resumen cladístico de lo que
se expone en el artículo principal puede
servir el árbol siguiente en el que sólo
aparecen taxones de distinto rango
entroncados directamente con la clase
Mammalia o pendientes de una jerarquización
más precisa: |
Los primeros mamíferos, los
cuales evolucionaron de los reptiles mamiferoides,
dieron su aparición en este período, posiblemente por
los cambios de clima que se hicieron y la deriva
continental, que motivaron la gran regresión marina del
Triásico: todas las tierras se unieron para formar el
supercontinente Pangea (dividido nuevamente a principios
del Jurásico, en la manera inversa de una transgresión
marina (por eso pasó a predominar los carbonatos).
Los sinápsidos (Synapsida,
gr. "arco fusionado"), o terópsidos (Theropsida) son una
clase de amniotas que incluye a los mamíferos y a todas
aquellas formas más relacionadas con ellos que con el
resto de amniotas. Los sinápsidos no mamíferos se han
denominado tradicionalmente "reptiles mamiferoides", es
decir reptiles similares a los mamíferos.
El rasgo craneano que los
caracteriza es la presencia de un orificio en los lados
del cráneo tras las órbitas de los ojos, la fosa
temporal inferior. Hace tiempo se supuso erróneamente
que esta fosa se había formado por la fusión de las dos
que existen en los reptiles diápsidos y por ello el
grupo recibió el inadecuado nombre de sinápsidos. El
estudio de los sinápsidos fósiles nos muestra cómo los
tipos más antiguos se parecen a los primeros reptiles y
los más avanzados son casi idénticos a los mamíferos.
Los
primeros mamíferos propiamente dichos.
En esta época había un grupo
de reptiles que poseían características de mamíferos;
eran muy frecuentes desde el periodo pérmico al triásico
y se conocen como el orden de los Terápsidos. Con el
tiempo, dieron lugar a distintas ramas evolutivas en las
que fueron apareciendo rasgos típicos de los mamíferos
al tiempo que se entremezclaban con los reptilianos.
Esto sucedió hace unos 180 millones de ańos cuando
declinaron dejando una línea de descendientes que no
resurgiría hasta pasados 100 millones de ańos, durante
los cuales los grandes reptiles predominaron en la
Tierra. Los primeros fósiles correspondientes de forma
definitiva a un mamífero fueron encontrados en rocas del
jurásico.
 |
Durante este periodo
existían ya cinco órdenes distintos de
mamíferos. Uno estaba compuesto por mamíferos
pequeńos similares a los roedores, que tenían
características dentales típicas de estos
animales, pero se extinguieron durante el
eoceno. Un segundo orden consistía en mamíferos
pequeńos y carnívoros, con molares de tres
cúspides y que también se extinguieron antes de
finalizar el eoceno. El tercer grupo estaba
formado por mamíferos del tamańo de una rata o
un topo; tenían un régimen alimentario
insectívoro y casi con toda seguridad los
podemos identificar como los antecesores de los
mamíferos actuales. |
Se desconoce con exactitud,
cuándo y dónde aparecieron los monotremas en la Tierra.
La mayor parte de los fósiles han sido encontrados en
Australia, si bien recientemente han sido descubiertos
en Argentina dientes fósiles de un ornitorrinco
perteneciente al género Monotrematum, lo
que sugiere que se distribuían por las regiones más
meridionales de Gondwana. El fósil más antiguo del que
se dispone de un monotrema data de más de 100 millones
de ańos. Se trata de una porción de la mandíbula de un
Steropodon galmani, un antepasado del
ornitorrinco que debía de ser del tamańo de un gato.
Los primeros metaterios eran
parecidos a las actuales zarigüeyas (Didelphimorphia) y
surgieron sobre las tierras de la actual Norteamérica
hace entre 80 y 100 millones de ańos. Desde allí se
extendieron por todos los continentes, pero fue al sur
donde proliferaron sin dificultad y a través de la unión
de estas tierras con el continente Australiano, lo
colonizaron poco antes de que ambos se separaran
definitivamente, quedando aislados del resto de los
mamíferos. Por su parte, los primeros euterios eran
similares a las actuales musarańas (Soricomorpha). Se
desconoce su origen cierto y durante algunos millones de
ańos tuvieron que convivir con los dinosaurios,
evolucionando lentamente hasta que éstos acabaron por
desaparecer y los mamíferos tuvieron que evolucionar
para colonizar la totalidad de la tierra, prácticamente
a su disposición.
El Cretáceo termina con la
desaparición de los dinosaurios hace alrededor de 65
millones de ańos, el clima frío dominante va
suavizándose y la supremacía de los primitivos
mamíferos, más evolucionados que los reptiles, con
capacidad para regular la temperatura corporal, un
sistema nervioso central más desarrollado y un organismo
preparado para adaptarse fácilmente a las condiciones
ambientales hace que éstos se diversifiquen y
evoluciones rápidamente para aprovechar los recursos de
un planeta que los dinosaurios al desaparecer han dejado
virtualmente a su disposición.
Bibliografía Sugerida.
Robert L. Carroll, Vertebrate
Paleontology and Evolution, W. H. Freeman and Company,
New York, 1988 ISBN 0-716-71822-7. Chapters XVII through
XXI.
Martin Prestia. 2002. żQUE ES
UN MAMIFERO?. Naturaleza.com.ar
Nicholas Hotton III, Paul D.
MacLean, Jan J. Roth, and E. Carol Roth, editors, The
Ecology and Biology of Mammal-like Reptiles, Smithsonian
Institution Press, Washington and London, 1986 ISBN
0-87474-524-1
T. S. Kemp, The Origin and
Evolution of Mammals, Oxford University Press, New York,
2005 ISBN 0-19-850760-7
Zofia Kielan-Jaworowska,
Richard L. Cifelli, and Zhe-Xi Luo, Mammals from the Age
of Dinosaurs: Origins, Evolution, and Structure,
Columbia University Press, New York, 2004 ISBN
0-231-11918-6.
Comprehensive coverage from
the first mammals up to the time of the K-T mass
extinction. Zhe-Xi Luo, "Transformation and
diversification in early mammal evolution", Nature
volume 450 number 7172 (13 December 2007) pages
1011–1019. doi:10.1038/nature06277. A survey article
with 98 references to the scientific literature.
Laurin, M. & Reisz, R. R.
2007. Synapsida: Mammals and their extinct relatives.
Version 06 April 2007.
The Tree of Life Web Project
Carroll, R. L. 1988.
Vertebrate Paleontology and Evolution. New York: W.H.
Freeman & Co.
Benton, M. J. 2005.
Vertebrate Paleontology, 3rd ed.
Oxford: Blackwell Science
Ltd.
Young, J. Z. 1977.
La vida de los vertebrados.
Editorial Omega, Barcelona, 660 pp.
Carroll, R.L. (1969).
"Problems of the origin of
reptiles." Biological Reviews, 44:
393-432.
Estudio y Campo de Investigación
de la Paleopalinología. |
|
paleopalinología
paleopalinología paleopalinología paleopalinología
paleopalinología paleopalinología paleopalinología |
|
Fragmento del Articulo:
Magnussen Saffer, Mariano. (2009). Estudio y Campo de
Investigación de la Paleopalinologia. Paleo, Boletín
Paleontológico. Ańo 7 numero 35: 19 - 21. |
|
paleopalinología
paleopalinología paleopalinología paleopalinología
paleopalinología paleopalinología paleopalinología |
La
paleopalinología se basa en la capacidad de polen y esporas
(también denominados palinomorfos) para ser fosilizados. Los
palinomorfos pueden ser transportados por el viento, o a través
de otros vectores como los animales, y depositados sobre
diferentes materiales. Estos microfósiles sedimentados a lo
largo del tiempo experimentan procesos de fosilización de su
exina de manera que se puede extraer, datar e identificar el
polen de un determinado material y deducir así cómo era la
vegetación en el pasado.
La palinología
es la disciplina que estudia polen, esporas, dinoflagelados y
cualquier palinomórfo actual o fósil. El estudio palinológico de
polen actual contribuye a la taxonomía de plantas, certificar
calidad de mieles, predecir cosechas e investigaciones
agronómicas, estudiar la naturaleza de los contaminantes
biológicos (polen alergénico) entre otras aplicaciones. Los
analisis palinologicos son tambien util a la Arqueologia,
Paleontología, Climatologia, Estratigrafía, Geología del
petróleo entre otras aplicaciones. Ésta se centra
fundamentalmente en el análisis de su morfología externa que
presenta patrones estructurales diferentes a tenor de las
variaciones en la exina, que es la pared externa de los granos
de polen. El estudio y análisis microscópico de su simetría,
aperturas en las paredes, contorno, forma, tamańo, etc. tiene un
valor taxonómico y permite distinguir taxones diferentes a
distintos niveles (familia, géneros, especies). Es en el estudio
paleontológico donde alcanza su máxima versatilidad, pues el
polen tiene gran resistencia a la putrefacción debido a las
características químicas de la exina. Esta área de investigación
se denomina "Paleopalinología".
|
 |
Este hecho tiene importancia biológica en
sí misma dado que de ello se pueden deducir multitud de
parámetros como la evolución vegetal, extinciones de
especies vegetales y todos aquellos aspectos
relacionados con la biogeografía histórica de las
plantas (geobotánica o fitogeografía). Además se pueden
inferir otras características del paisaje que en
conjunción con otras disciplinas (arqueozoología,
paleoantropología, paleobotánica) permite la
reconstrucción de ambientes fósiles en distintos planos.
<<<< Investigador
observando polen por un microscopio. |
Los granos de
polen y las esporas son microscópicos pero cada cual tiene una
forma peculiar a su especie. Están constituidos por una membrana
exterior (exina) muy resistente que les permite soportar altas
temperaturas y presiones. Son resistentes a la degradación
biológica y química y preservan sus estructuras. Quedan
momificados en el hielo, en las turbas, en las resinas fósiles
(ámbar) y en rocas sedimentarias que tienen millones de ańos.
Están bien conservados pero no son viables. Son útiles a la
geología del petróleo y del carbón y a la estratigrafía ya que
permite efectuar correlaciones regionales.
Entre los
microfósiles existe un gran número de especies diferentes,
pudiéndose encontrar polen producido por las gimnospermas y las
angiospermas, esporas producidas por los helechos y hongos y
microorganismos acuáticos. Proporcionan una información muy
completa de la vegetación existente, ya que usualmente se
encuentran muchas formas diferentes de polen y esporas.
Adicionalmente la palinología permiten datar las rocas y conocer
la evolución y las asociaciones de los diferentes grupos
vegetales. También proporciona información sobre la naturaleza
de los sedimentos, cantidad de materia orgánica y conocer si los
depósitos son marinos o continentales
|
 |
De hecho la paleoclimatología emplea la
paleopalinología como herramienta frecuentemente. Este
uso se debe a que las especies vegetales mantienen
óptimos ambientales bajo distintas circunstancias
climáticas. Una simplificación del mecanismo inductivo
que se emplea para la reconstrucción climática es el
siguiente: si una especie que hoy es típica de ambiente
frío y húmedo es hallada depositada en materiales de una
determinada época del pasado podemos inferir que el
clima durante ese periodo era frío y húmedo.
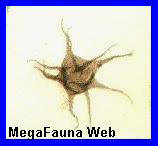
<<<<
Grano de polen (Multiplicisphaeridium) visto al
microscopio |
Por supuesto el
proceso de interpretación de la ecología de ambientes pasados no
es tan simple y se deben considerar numerosos aspectos como la
interacción entre las diferentes especies dentro de una
comunidad vegetal, la amplitud ecológica de los taxones, etc.
Algunas de las
fuentes de polen fósil más frecuentes, especialmente en
latitudes medias del planeta, son las turberas, ambientes
lacustres o humedales. La conservación del polen pasa por
mantener condiciones de anoxia de manera constante, de modo que
la preservación óptima del polen se da en ambientes poco
expuestos a la erosión y oxigenación. Por ello, las turberas y
demás espacios lacustres se muestran especialmente útiles en
este aspecto.
|
 |
Por otra parte, en este tipo de cuencas
de depósito el material sedimentado lo hace de manera
secuencial de modo que se pueden extraer sondeos
coherentes en el tiempo, en los que la base sea más
antigua que la zona superior. Si se realizan análisis
polínicos de muestras consecuentes en un sondeo de estas
características y además se realizan dataciones
absolutas se pueden obtener curvas de cambio vegetal a
lo largo de un periodo concreto y por tanto secuencias
de cambio climático a lo largo de dicho periodo.
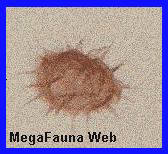
<<<<<
Grano de polen Multiplicisphaeridium sp
visto al
microscopio |
La
paleopalinología se ha convertido en una disciplina que sirve de
gran apoyo a otras áreas de estudio paleontológicas para la
deducción de resultados paleoambientales. Es también usada en
arqueología de la misma forma.
Para estudiar el
polen y las esporas, se tritura la roca y luego se ataca con
ácidos que suprimen los minerales y permite aislar la materia
orgánica y los microfósiles. Ácido clorhídrico para atacar los
carbonatos, ácido fluorhídrico para la disolución de la sílice,
ácido nítrico para oxidar y eliminar el exceso de materia
orgánica, blanquear y limpiar el polen y las esporas que se van
estudiar. Los estudios se realizan con microscopio óptico o en
microscopio electrónico de barrido.
Por comparación
con especies actuales y fósiles, se les clasifica en familias,
género y especies. Para indicar que es polen fósil se les
aproxima a la especie actual se agrega el sufijo dites .
Ejemplos: Proteacidites,
Podocarpidites,
Nothofagidites. A las esporas en cambio se
les agrega generalmente el sufijo sporites. Si no hay afinidad
conocida se crean géneros y especies artificiales.
Fuente Sugerida:
ARCHANGELSKI,
S. (1970). Fundamentos de Paleobotánica. Universidad Nacional de
La Plata.
Serie Técnica y Didáctica nş 11.
BOUREAU, E. Traité de Paléobotanique.
MassoN. (4 vols)
DARRAH, W.C. Principles of Paleobotany. Roland Press Co.
DILCHER, D. & TAYLOR, T.N. Eds. (1980). Biostratigraphy of
fossil plants.
Dowden,
Hutchinson& Ross, Inc.
EMBERGER, L. (1968). Les plants fossiles. Les Vegetaux vivants.
Masson et Cie.
FONT QUER, P. Diccionario de Botánica. Sa Reimpresión. Editorial
Labor.
GIFFORD, E. M. & FOSTER, A. Morpholiogy and
Evolution of Vascular Plants.
Freeman & Co.
3rd ed.
MORET, L. (1964). Manuel de Paléontologie Végetale.
Masson.
RIDING, R. Ed. (1991). Calcareous Algae and Stromatolites.
Springer-Verlag.
SAENZ DE RIVAS, C. (1978). Polen y esporas. Ediciones Blume.
SCOT, D.H. (1963).
Studes in Fossil Botany. Haffner Publishing Company. (4 Vols.).
STASBURGER, E. Tratado de Botánica.
8Ş Edición
espańola. Ediciones Omega S.A.
TRAVERSE, A. (1988). Paleopalynology. Unwin Hyman Ltd.
Cope y Marsh. Amigos, enemigos y la guerra de los
huesos a fines del siglo XIX.
Tomado de;
Magnussen Saffer,
Mariano (2010). Cope y Marsh.
Amigos, enemigos y la guerra de los huesos a fines del siglo
XIX.
Paleo, Boletín
Paleontológico. Ańo 8. 46: 19-22.
marianomagnussen@yahoo.com.ar
Othniel Charles Marsh y Edward
Drinker Cope fueron dos de los grandes pioneros de la
paleontología en Estados Unidos y sus carreras científicas
poseen entidad de sobra para haberles hecho famosos
individualmente. Pero es por su enemistad por lo que son
universalmente recordados. En los números 044 y 045 de Paleo -
Boletín Paleontológico he publicado la biografía individual de
Othniel Charles Marsh y Edward Drinker Cope, quedando pendiente
la mencionada “Guerra de los Huesos”, que los convirtieron en
grandes figuras mediáticas de la paleontología americana.
Cope y Marsh eran ambiciosos,
arrogantes y vengativos, sedientos de fama y renombre. Los
corroyeron los celos, la envidia y la desconfianza. La
competencia sacó a relucir lo peor de ambos. Cope llegó a
escribir que le gustaría ver a Marsh empalado en los cuernos de
un Monoclonius sphenocerus, lo que, hay que
convenir, resulta rebuscado (y doloroso). También pidió que tras
su muerte le extrajeran el cerebro y lo pesaran para compararlo
con el de Marsh. Por su parte, Marsh calificó a su rival de
´histérico y le sugirió que se volase la tapa de los sesos.
 |
Deseaban los dos no sólo
vencer en la carrera por encontrar y describir nuevos y
más grandes dinosaurios, sino también que el otro
fracasara. Ambos perdieron a sus madres de nińos, a los
tres ańos. Cope, impaciente y temperamental,
pendenciero, era más brillante; Marsh, más meticuloso y
fiable, y más influyente. De familias ricas, los dos
disfrutaron de situaciones acomodadas y buenas rentas,
que dedicaron a conseguir fósiles y a labrarse una
carrera científica. Se conocieron en Berlín en 1864,
cuando viajaban por Europa para profundizar en sus
conocimientos científicos (y de paso librarse de ser
movilizados en la guerra de secesión). Pasaron un par de
días de manera bastante amistosa y luego, ya en casa, se
escribieron y visitaron algunos yacimientos juntos.
Hasta se dedicaron especies. |
No está claro cómo empezaron a
odiarse. Parece que Marsh trató de comprar unos fósiles
apalabrados a Cope por el propietario de unos terrenos ricos en
ellos.
or su parte, Cope invadió una
zona de caza de Marsh en Kansas. En 1870, Marsh examinó un
esqueleto de Elasmosaurus (un plesiosaurio)
montado por Cope y dijo que estaba al revés: la cabeza en la
cola. Cope, lógicamente ultrajado, se puso furioso. Pero Marsh
tenía razón. Él mismo, sin embargo, patinó en otras ocasiones,
equivocando las cabezas de dos saurópodos o bautizando a un
mismo bicho con dos nombres diferentes. La Guerra de los Huesos
fue un periodo de intensa especulación y descubrimientos de
fósiles durante la Gilded Age de la historia de los Estados
Unidos. Los dos paleontólogos utilizaron métodos deshonestos
para superar al otro en el campo, recurriendo a sobornos, robos,
y destrucción de huesos. Los científicos también se atacaron
mutuamente en obras científicas, intentando arruinar la
credibilidad del otro y dejarlo sin financiación.
Su investigación de huesos los
llevó al oeste, a los ricos yacimientos paleontológicos de
Colorado, Nebraska y Wyoming. Entre 1877 y 1892, ambos
utilizaron su patrimonio y su influencia para financiar sus
propias expediciones y obtener servicios y fósiles de cazadores
de dinosaurios. Al final de la Guerra de los Huesos, ambos
habían agotado su patrimonio para alimentar su intensa
rivalidad.
Cope y Marsh tenían orígenes
muy diferentes. Cope nació en el seno de una familia cuáquera,
rica e influyente de Filadelfia. Aunque su padre quería que Cope
trabajara de granjero, este se distinguió como naturalista. En
1864, Cope se convirtió en profesor de zoología en el Haverford
College y se unió a Ferdinand Hayden en sus expediciones en el
oeste. Marsh, hijo de una familia de pocos medios de Lockport,
Nueva York, habría crecido en la pobreza de no haber sido por la
ayuda de su tío, el filántropo George Peabody. Marsh convenció a
su tío para que construyera el Museo Peabody de Historia
Natural, y consiguió el cargo de director del museo. Eso, junto
con la herencia que recibió de Peabody cuando este murió en
1869, hizo posible que Marsh disfrutara de una cómoda posición
financiera aunque, debido en parte a la visión estricta que
tenía Peabody del matrimonio, Marsh permanecería soltero toda la
vida.
 |
En una ocasión, los dos
científicos se habían marchado a una expedición de
recolección de fósiles en los pozos de marga de Cope en
Nueva Jersey, donde William Parker Foulke había
descubierto el holotipo del dinosaurio Hadrosaurus
foulkii, descrito por el paleontólogo Joseph
Leidy; este fue uno de los primeros descubrimientos de
dinosaurios en América, y los pozos todavía eran ricos
en fósiles. Aunque los dos fueron amigablemente, Marsh
sobornó en secreto a los operadores de los pozos para
que le llevarán los fósiles que descubrían a él, en
lugar de a Cope. Los dos empezaron a atacarse mutuamente
en documentos y publicaciones, y su relación personal
empeoró. |
En los artículos de prensa,
Cope atacó a Marsh por plagio y mala gestión financiera, y atacó
a Powell por sus errores de clasificación geológica y por
malversar el dinero otorgado por el gobierno. Marsh y Powell
pudieron publicar su visión del asunto, lanzando sus propias
acusaciones contra Cope. Los artículos de Ballou estaban poco
estudiados, escritos y leídos, y el propio Cope escarmentó tras
un artículo de The Philadelphia Inquirer que sugería que los
mandatarios de la Universidad de Pensilvania pedirían la
dimisión de Cope a menos que presentara pruebas para sus
acusaciones contra Marsh y Powell.
El mismo Marsh mantuvo viva la
historia del Herald con una feroz réplica, pero a finales
de enero la historia desapareció de los diarios, y poco había
cambiado entre los rivales acérrimos. No se celebró ningún
juicio del congreso para investigar la malversación de fondos de
Powell y ni Cope ni Marsh hubieron de responder sobre su
responsabilidad en los errores, pero parte de las calumnias de
Ballou contra Marsh quedaron asociadas con el Servicio. Ante un
sentimiento antiservicio enardecido por la sequía en el oeste y
la preocupación sobre la apropiación de las granjas abandonadas
en la misma zona, Powell se encontró bajo una estrecha
vigilancia por parte del House Appropriations Committee.
|

|
Llevado a actuar por el
supuesto derroche de Marsh en cuanto a los fondos del
Servicio, el Appropriations Committee exigió que el
presupuesto del Servicio fuera detallado. Cuando se puso
fin a su apropiación en 1892, Powell envió un lacónico
telegrama a Marsh indicándole que esperaba su dimisión,
una afrenta personal además de financiera. Al mismo
tiempo, muchos de los aliados de Marsh se estaban
retirando o habían muerto, reduciendo su crédito
científico. Justo cuando el extravagante estilo de vida
de Marsh empezaba a pasarle factura, Cope consiguió un
cargo en el Servicio Geológico de Texas, aunque, todavía
tambaleándose por los ataques personales que había
recibido durante el asunto del Herald, no
continuó con los suyos. |
La suerte de Cope mejoró
durante la primera parte de la década de 1890, cuando fue
promovido en el lugar de Leidy como Profesor de Zoología y fue
elegido presidente del American Association for the Advancement
of Science el mismo ańo que Marsh dejó su cargo al frente de la
Academia de Ciencias. Sin embargo, a finales de la década su
suerte volvió a empeorar, a medida que Marsh recuperaba parte de
su prestigio, consiguiendo la Medalla Cuvier, el galardón
paleontológico más importante.
Marsh permaneció soltero, y en
la profesión corría el chascarrillo de que manejaba muchos
huesos, pero nunca consiguió una costilla. Cope se casó y tuvo
una hija. Parece haber sido un donjuán, y de hecho hay quien ha
atribuido su muerte a la sífilis, contraída durante sus
expediciones paleontológicas (´Ninguna mujer estaba a salvo de
Cope a menos de ocho millas , dijo de él un amigo). Otros, sin
embargo, atribuyen su fallecimiento a que se automedicaba los
problemas gastrointestinales ingiriendo el formaldehído de
preservar especímenes.
La herencia de los dos
paleontólogos es enorme. Cope y Marsh quedaron económica y
socialmente arruinados por sus esfuerzos para deshonrar al
contrario, pero sus contribuciones a la ciencia y la disciplina
de la paleontología serían inmensas; a su muerte, los
científicos dejaron toneladas de cajas de fósiles sin abrir.
Entre ambos descubrieron y documentaron más de 130 nuevas
especies de dinosaurio, incluidos los tan populares
Diplodocus, Stegosaurus, Allosaurus, Camarasaurus, Coelophysis
y Triceratops. Lograron esqueletos
completos que montados en posiciones dramáticas en los grandes
museos cautivaron la imaginación de las multitudes. Fueron
pioneros en sistemas de excavación y conservación. Así que puede
decirse que la suya fue una rivalidad muy productiva para la
ciencia. Aunque, desde luego, no un gran ejemplo.
Bibliografía Sugerida.
Baalke, Ron (13-10-1994). "Edward
Cope's Skull". Lista de correo lepomis.psych.upenn.edu.
Consultado el 15 de agosto de 2008.
Jaton, Jacinto. (2008). Ese
diplodocus es mío. Publicado en El Pais.
Norell, Mark A.; Gaffney, Eric
S.; i Dingus, Lowell (1995). Discovering Dinosaurs in the
American Museum of Natural History. Nova York: Alfred Knopf
Publishing
Preston, Douglas (1993).
Dinosaurs in the Attic: An Excursion Into The American Museum of
Natural History. Estados Unidos: Macmillan Publishers
Sterling, Keir (1997).
Biographical Dictionary of American and Canadian Naturalists and
Environmentalists. Greenwood Publishing Group
Wallace, David Rains (1999).
The Bonehunters' Revenge: Dinosaurs, Greed, and the Greatest
Scientific Feud of the Gilded Age. Houghton Mifflin Books.
El Súper Árbol de los
Dinosaurios.
Durante mucho
tiempo se ha debatido si los dinosaurios eran parte de la
'Revolución terrenal "que se produjeron unos 100 millones de
ańos durante el Cretácico cuando las aves, mamíferos, plantas
con flores, insectos y reptiles todos se sometió a una rápida
expansión.
Un estudio
internacional, dirigido por la Universidad de Bristol, muestra
que durante los últimos 50 millones de ańos de existencia, los
dinosaurios no fueron la ampliación de tan activa como se había
pensado previamente y que la aparente explosión de la diversidad
de dinosaurios se puede explicar en gran parte por el sesgo de
muestreo.
El equipo elaboró un 'supertree' de los dinosaurios, es el más
probable patrón de evolución de 440 de las 600 especies
conocidas de dinosaurios. "Supertrees son muy grandes árboles de
la familia hizo uso de sofisticadas técnicas de ordenador que
cuidadosamente puntada juntos varios pequeńos árboles que fueron
previamente elaborados por expertos en los diversos subgrupos",
explicó el autor principal Graeme Lloyd.
 |
"Nuestro supertree resume los esfuerzos de dos décadas
de investigación de cientos de trabajadores de
dinosaurios en todo el mundo y permite a buscar todas
las pautas inusuales en el conjunto de los dinosaurios
por primera vez." Es la imagen más completa hasta la
fecha de cómo evolucionaron los dinosaurios . Los
resultados se publican hoy (23 de julio) en las
labores de la Royal Society B. Mike Benton Profesor
de la Universidad de Bristol, dijo: "No es completa,
pero es la más detallada y completa único árbol
evolutivo de los dinosaurios producido, y, de hecho,
para casi cualquier otro grupo. |
"Hasta ahora,
la mayoría de los estudios de la evolución de los dinosaurios no
fueron probados en contra de una numéricamente precisa y
completa base de datos. Esperamos que nuestro estudio marca la
primera de una nueva ola de este tipo en profundidad, los
estudios cuantitativos en paleontología. "
El nuevo
estudio utiliza técnicas estadísticas para distinguir
inusualmente altas tasas de diversificació n de las tasas
normales. Los resultados muestran que todas las ráfagas de
diversificació n que ocurrió en los primeros cincuenta millones
de ańos de evolución de los dinosaurios.
 |
Más
tarde expansiones no se distinguen de las tasas
normales. Esto sugiere dinosaurios no aprovechar las
ventajas del nuevo suministro de alimentos disponibles
durante el Cretácico Terrestre Revolución - tales como
plantas con flores, lagartos, serpientes, aves y
mamíferos.
El
trabajo fue realizado utilizando la Computación de Altas
Prestaciones instalaciones de la Universidad Nacional de
Irlanda, Maynooth. Se basa en una combinación de 155
publicado dinosaurio 'árboles' y tomó aproximadamente
5000 horas de tiempo de cálculo. |
La atención se
centró clave para ver si los dinosaurios ha sido parte de una
importante fase de evolución en la tierra - la Revolución
Terrestre Cretácico (entre 125-80 millones de ańos atrás) -
cuando muchos nuevos grupos de plantas y animales se expandió
rápidamente. Durante este tiempo, las plantas con flores y los
insectos sociales surgieron y se convirtió cada vez más común.
Muchos animales también backboned ampliado para aprovechar las
nuevas fuentes de alimentos.
Para conocer
más y bajar el árbol genealógico de los dinosaurios en PDF,
súmese gratuitamente a nuestro
grupo de correo
Web.
El desarrollo de la filosofía natural
y la paleontología hasta el siglo XVII.
Por Mariano
Magnussen Saffer. Grupo Paleo. Museo Municipal Punta Hermengo de
Miramar.
Fundación Argentavis.
marianomagnussen@yahoo.com.ar. Tomado de; -
Magnussen Saffer,
Mariano. 2015. Paleo Revista Argentina de
Paleontología.
El desarrollo de la filosofía natural
y la paleontología hasta el siglo XVII.
Boletín Paleontológico. Ańo XIII. 135:
20-22.
La historia de
la paleontología recorre la historia de los esfuerzos para
entender la historia de la vida en la Tierra a través del
estudio del registro fósil dejado por organismos vivos. Ya que
tiene que ver con la comprensión de los organismos vivos del
pasado, la paleontología es considerada como un campo de la
biología, pero su desarrollo histórico ha estado estrechamente
ligado a la geología y el esfuerzo para entender la historia de
la Tierra misma.
Jenófanes de
Colofón, un filósofo griego, que vivió en el siglo sexto antes
de Cristo (570-480 a. C.), reconoció que algunas conchas fósiles
eran restos de moluscos, lo que utilizó como evidencia para
argumentar que lo que en el momento era tierra firme alguna vez
había estado bajo el mar.
Leonardo da
Vinci (1452-1519), mucho mas tarde, en un cuaderno inédito,
también llegó a la conclusión de que algunas conchas marinas
fósiles eran restos de moluscos. Sin embargo, en ambos casos,
los fósiles eran restos completos de especies de moluscos que se
parecían mucho a las especies vivas, y por lo tanto eran fáciles
de clasificar.
 |
En
1027, el naturalista persa Ibn Sina (conocido como
Avicena en Europa), propuso una explicación de cómo
se producía la pedregosidad de los fósiles en El
libro de la curación. Modificó una idea de
Aristóteles, que explicó este proceso en términos de
exhalaciones vaporosas. Ibn Sina modificó esta idea en
su teoría de los fluidos petrificantes (succus
lapidificatus), la cual fue retomada y extendida por
Alberto de Sajonia en el siglo
xiv y era
aceptada en alguna forma por la mayoría de los
naturalistas del siglo
xvi. |
Shen Kuo
(1031-1095) un chino de la dinastía Song utilizó fósiles marinos
encontrados en las montańas Taihang para inferir la existencia
de procesos geológicos como la geomorfología y las fluctuaciones
de la orilla del mar con el tiempo. Haciendo observaciones de
bambúes petrificados preservados bajo tierra en Yan'an, región
de Shanbei, provincia de Shaanxi, formuló una teoría del cambio
climático gradual, ya que Shaanxi era parte de una zona de clima
seco que no proporcionaba un hábitat adecuado para el
crecimiento del bambú.
Como resultado
del nuevo énfasis en la observación, clasificación y
catalogación de la naturaleza, los filósofos naturales del siglo
xvi en Europa
comenzaron a establecer extensas colecciones de objetos fósiles
(así como colecciones de especímenes de plantas y animales), que
a menudo se almacenaban en gabinetes especialmente construidos a
su apropiada organización.
Conrad Gesner
publicó en 1565 un trabajo acerca de los fósiles que contenía
una de las primeras descripciones detalladas de un gabinete y
colección. La colección pertenecía a uno de los miembros de la
extensa red de corresponsales que de la que Gesner obtenía
información para sus trabajos. Este tipo de redes de
correspondencia informales entre filósofos naturales y
coleccionistas se volvió cada vez más importante en el
transcurso del siglo xvi
y fue precursor directo de las sociedades científicas que
comenzarían a formarse en el siglo
xvii. Estas
colecciones en gabinete y redes de correspondencia jugaron un
papel importante en el desarrollo de la filosofía natural.
 |
Sin
embargo, la mayoría de los europeos del siglo
xvi no
reconocieron que los fósiles eran los restos de
organismos vivos. La etimología de la palabra fósil
proviene del latín para desentarrar, desenterrado.
En consecuencia, el término se aplica a una amplia
variedad de objetos pétreos sin tener en cuenta si
podrían tener un origen orgánico. |
Escritores del
siglo xvi como
Gesner y Georg Agricola estaban más interesados en la
clasificación de este tipo de objetos por sus propiedades
físicas y místicas que en la determinación de su origen. Además,
la filosofía natural del período animó a que se formularan
explicaciones alternativas para el origen de los fósiles. Tanto
las escuelas aristotélicas y neoplatónicas proporcionaron apoyo
a la idea de que los objetos pétreos pueden crecer dentro de la
tierra para parecerse a los seres vivos.
La filosofía
neoplatónica sostuvo que podía haber afinidades entre los
objetos vivos y no vivos que podrían causar que unos se
parecieran otros. La escuela aristotélica sostuvo que las
semillas de organismos vivos podrían introducirse bajo tierra y
generar objetos parecidos a esos organismos.
Bibliografía sugerida.
Bowler, Peter J.
(2003).
Evolution:The History of an Idea. University of California
Press.
Bowler, Peter J.; Iwan Rhys Morus (2005). Making Modern
Science. The University of Chicago Press.
Cadbury, Deborah (2000), The Dinosaur Hunters: A True Story
of Scientific Rivalry and the Discovery of the Prehistoric World,
Fourth Estate,
Desmond, Adrian (1975). "The Discovery of Marine Transgressions
and the Explanation of Fossils in Antiquity". American Journal
of Science, Volume 275.
Palmer, Douglas (2005) Earth Time: Exploring the Deep Past from
Victorian England to the Grand Canyon. Wiley, Chichester. I
Rudwick, Martin J.S. (1985). The Great Devonian Controversy: The
Shaping of Scientific Knowledge among Gentlemanly Specialists.
The University of Chicago Press.
Zittel, Karl Alfred von (1901). History of geology and
palaentology to the end of the Nineteenth Century.
Charles
Scribner's Sons, London.
Wikipedia. |













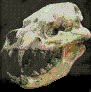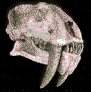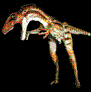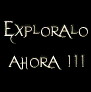

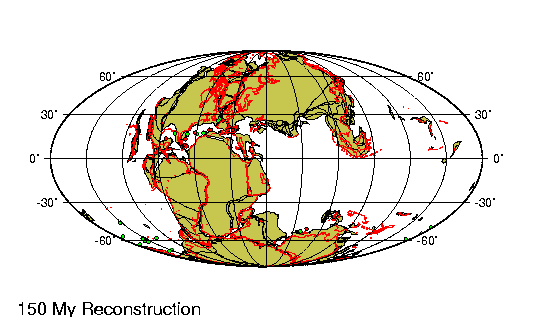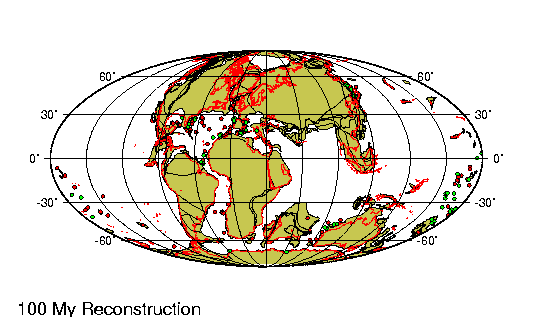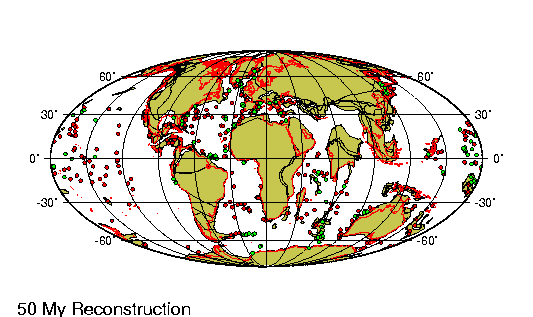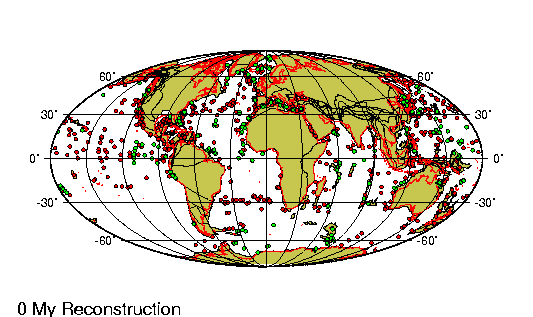


















{kind=link}