El Oligoceno es la tercera división
del periodo terciario del cenozoico, que se inició hace unos 35,4 millones de
ańos y finalizó hace unos 23,3 millones de ańos. Al igual que el eoceno, que lo
precedió, y que el mioceno, que vino a continuación, el oligoceno (del griego,
'poca vida') fue definido en función del porcentaje de especies modernas de
moluscos y crustáceos (10-15%) presentes en los estratos correspondientes a esta
era.
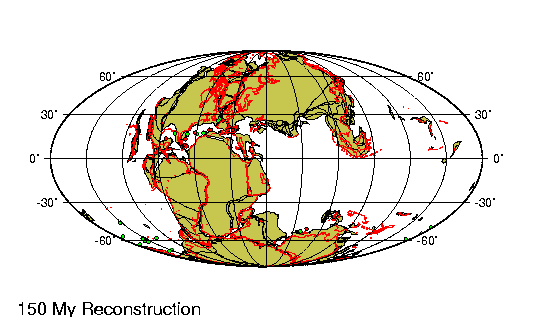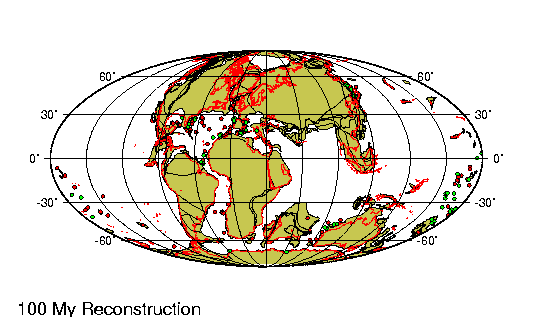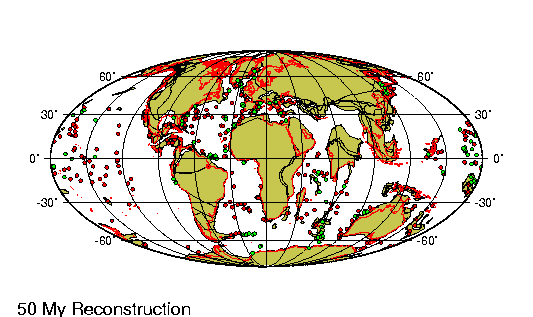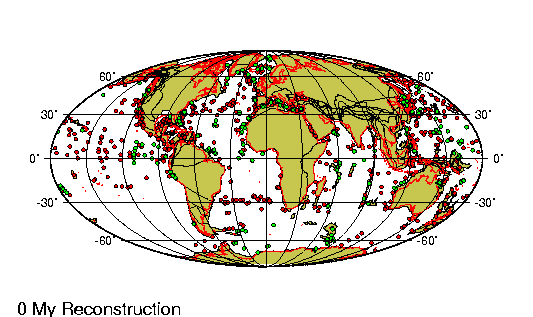
Según la teoría de la
tectónica de placas, las colisiones entre las placas de la corteza
terrestre continuaron sin pausa desde el eoceno. En el hemisferio
oriental, los restos afroárabes e indios del anterior
supercontinente de Gondwana chocaron con Eurasia al norte, cerrando
el extremo oriental del mar de Tetis y dejando en su lugar un
residuo muy mermado, el Mediterráneo. Las fuerzas de compresión
generadas por la colisión contribuyeron a elevar un extenso sistema
de cadenas de montańas, desde los Alpes en el Oeste hasta el
Himalaya en el Este.
Mientras tanto, la placa
australiana chocaba contra la indonesia, y la norteamericana había empezado a solaparse
sobre la del Pacífico. Como resultado, el proceso de expansión de suelo marino originado en
la dorsal del Pacífico oriental se desvió a una dirección
perpendicular al eje de la dorsal. Se produjo una gran falla —la
famosa falla de San Andrés, en San Francisco, generadora de
terremotos— para compensar este cambio en el movimiento entre
placas. El clima siguió siendo subtropical y húmedo en toda
Norteamérica y Europa, pero había comenzado una tendencia al
enfriamiento global a largo plazo, que culminaría en los periodos
glaciales del pleistoceno.
Nebraska,
Dakota del Sur(USA).
Los camellos de
ahora son un pequeńo grupo, que incluye los de una y dos
jorobas de África y Asia, así como a las llamas y vicuńas
de América del Sur. En el Mioceno, entre hace 25 y 5
millones de ańos, eran un exitoso grupo extendido por todo
el mundo. Uno de los camellos más antiguos fue el
Poebrotherium, un animal del tamańo de una
cabra, con patas largas y esbeltas y cuello largo, como en
las formas actuales. Las patas largas y los pies pequeńos
muestran que el Poebrotherium era un veloz
corredor. Los pies tenían pequeńos cascos en los dos dedos,
pero tales cascos se perdieron en la evolución del camello
y fueron sustituidos por anchos cojinetes, característica
que les permite caminar en la arena blanda. El
Poebrolherium tenía mandíbulas largas y los dientes
frontales sobresalían un poco, como en los camellos
modernos, lo que les permitía morder plantas en lugares muy
cercanos a la raíz.
América del Norte (Wyoming).
Tenia 14 cm de longitud, 37 cm de envergadura. Icaronycteris debió de ser casi idéntico a los
micromurciélagos modernos, pero todavía conservaba algunos
rasgos muy primitivos. Las alas eran relativamente cortas y
anchas, y la boca incluía una gran cantidad de dientes,
distribuidos como los de los insectívoros. No tenía el
cuerpo tan rígido como los murciélagos actuales y la cola
era larga y no estaba conectada con las patas traseras por
medio de una membrana de piel. Tanto el pulgar como el
primer dedo estaban provistos de una garra (mientras que los
murciélagos actuales sólo tienen garras en el pulgar), para
poder colgarse verticalmente de las paredes de las cavernas
o de otros soportes. Incluso en una etapa tan primitiva de
su evolución, los murciélagos descansaban cabeza abajo. Es
indudable que Icaronycteris vivía igual que
los micromurciélagos modernos, atrapando insectos con las
alas, probablemente mientras volaba cerca del agua al
atardecer, cuando había pocas aves por allí. Se han
encontrado algunos murciélagos en un estado de conservación
sorprendente, en los depósitos de esquistos petrolíferos de
Messel, cerca de Frankfurt-am-Mein, en Alemania. Incluso se
pueden ver las membranas de las alas, y los restos que se
han encontrado dentro del estómago del murciélago confirman
que se trataba de un insectívoro.
América del Norte (Nebraska).
Tenia 80 cm de longitud Hesperocyon, un animal
pequeńo y activo, con aspecto de mangosta o civeta, fue uno
de los miembros más primitivos de la familia de los cánidos.
Es posible que no se pareciera demasiado a un perro, con el
cuerpo y la cola largos y flexibles, las patas cortas y
débiles y los pies abiertos, dotados de cinco dedos. Sin
embargo, la estructura de los huesecillos del oído y la
distribución de sus dientes demuestran, sin ninguna duda,
que se trataba de un cánido primitivo. Los fósiles del
cráneo de esta criatura indican que las partes del oído
interno estaban encerradas en hueso, y no en un cartílago;
ésta es una característica propia de los perros, que
diferencia a Hesperocyon de otros carnívoros
más primitivos. Los dientes de Hesperocyon
demuestran que le faltaba el último molar a cada lado de la
mandíbula superior, con lo cual se totalizan 42 dientes, en
lugar de los 44 habituales. El último premolar superior y el
primer molar inferior de cada lado estaban modificados en
forma de cuchillas para cortar carne, típicas de los perros
y de la mayor parte de los carnívoros auténticos (es decir,
los que se incluyen dentro del orden de los carnívoros).
América del Norte (Nebraska).
Tenia 1 m de longitud. Cynodesmus fue uno de
los primeros cánidos que realmente se parecía a los perros
actuales. Tenía más o menos el tamańo y la forma de un
coyote, Canis latrans, como los que se
encuentran en la actualidad en América del Norte y América
Central. La cara, sin embargo, era más corta (el hocico
largo, propio de los perros, llegaría en una etapa muy
posterior de su evolución), y tenía el cuerpo bastante
largo, con una cola pesada. Las patas de Cynodesmus
eran semejantes a las de los perros, pero no corrían tan
bien como las de los perros actuales. Las praderas abiertas
de América del Norte no se habían formado todavía: su
aparición estimuló la evolución de mamíferos de movimientos
veloces que se alimentaban de hierba, lo cual produjo, a su
vez, la inmediata evolución de los cazadores de pies ágiles,
como los perros. Todavía había cinco dedos en cada una de
las patas de Cynodesmus, si bien los primeros
eran más cortos que los demás. Las garras eran estrechas y
retráctiles en parte, como las de los gatos, en vez de las
estructuras gruesas, romas y resistentes que se
desarrollarían en los perros posteriores. Es probable que Cynodesmus tuviera la costumbre de emboscar a sus
presas, al estilo gatuno, en lugar de perseguirlas y
echárseles encima, como hacen los perros.
América del Norte. Tenia 60 cm de altura en la
cruz. En los lugares donde los bosques cedían paso a zonas
más abiertas, los caballos dejaron de estar confinados a
retozar por el bosque bajo y comenzaron a desarrollar la
capacidad de trotar y de galopar. Mesohippus
tenía un tamańo similar al del galgo y el cuerpo medía 1.2 m
de largo; era algo más grande que sus predecesores, pero las
patas, provistas de tres dedos, eran más ágiles. El dedo
central era más grande que los demás. Los premolares
comenzaban a parecerse a los molares, lo cual aumentaba la
superficie de masticación y, por ende, su eficacia, aunque
las coronas seguían siendo bajas. Estos dientes requerían
una mandíbula poco profunda, por lo que la cabeza era
bastante larga y puntiaguda.
Dakota del Sur (USA), Europa y Asia. Los
brontoterios fueron un grupo de grandes herbívoros que se
parecían mucho a los rinocerontes, pero que sólo estaban
lejanamente emparentados con ellos. El Brontotherium
y sus parientes se alimentaban de hojas de árboles y
arbustos, y generalmente vivían en los bosques templados que
cubrían América del Norte, Europa y partes de Asia. El Brontotherium era más grande que cualquier
rinoceronte actual, y tenía en el hocico un notable "cuerno"
bifurcado cuya forma recuerda una catapulta. Tal "cuerno"
era mayor en los machos que en las hembras; tal vez servía
para peleas, como los cuernos de los ciervos y antílopes de
ahora. Los brontoterios se extinguieron cuando los bosques
fueron reemplazados por grandes pastizales. No podían
alimentarse de los nuevos pastos, y su lugar lo ocuparon
caballos, rinocerontes y otros animales que sí podían
hacerlo.
América del Norte, Usa. Tenia 3 metros de
longitud. Daeodon era un gigante mamífero que
se alimentaba de vegetales, carrońa y de animales pequeńos.
Vivía en praderas y bosques abiertos. Estos animales
grandes, parecidos a los cerdos, que probablemente tuvieron
su origen en Asia a fines del Eoceno, se hicieron frecuentes
en Europa y Asia y se dispersaron hacia América del Norte.
Tuvieron su etapa de mayor proliferación durante el
Oligoceno, pero algunos sobrevivieron en América del Norte
hasta el comienzo del Mioceno, hace unos 20 millones de
ańos. Algunos eran inmensos, llegando a alcanzar las
proporciones de un hipopótamo. Una característica destacada
de los entelodóntidos, como Daeodon era la
presencia de dos pares de prominencias óseas que sobresalían
de los costados de la mandíbula inferior.
América del Norte (Dakota del Sur). Tenia 1 m de
longitud Este animal pequeńo y gracioso, semejante al
ciervo, vivía en los bosques de las tierras altas del oeste
de América del Norte. Era uno de los primeros miembros de la
familia y todavía era bastante primitivo, por cuanto tenía
cuatro dedos en las patas delanteras y las traseras. La
característica mas notable de Protoceras y, en
realidad, de todos los protocerátidos, y la que mas los
distingue de los ciervos, era la disposición de Ios cuernos
en la cara. A diferencia de la ornamentación del ciervo, no
se trataba de astas, ni se mudaban cada ańo. tampoco es
correcto llamarlos «cuernos», puesto que no estaban
recubiertos de esta sustancia. En realidad, eran
protuberancias -óseas que, probablemente estaban cubiertas
de piel, como los caronas de las jirafas. Es evidente que la
disposición de los cuernos respondía a algún tipo de
estructura de adorno que se utilizaba para el acoplamiento o
para alejar a los rivales. Es probable que las prominencias
fueran más efectivas vistas desde los lados que desde el
frente. Los primeros protocerátidos conservaban los
incisivos superiores en la parte anterior. Sin embargo,
cuando apareció Protoceras, éstos se habían
perdido, de modo que los dientes inferiores funcionaban
contra una callosidad ósea situada en la mandíbula superior,
igual que en los ciervos y los bóvidos actuales.
América del Norte. Tenia 2,5 m de altura en la
cruz. Cuando el Eoceno se convirtió en Oligoceno, los
brontoterios se volvieron enormes, más grandes que
cualquiera dé los rinocerontes vivos, y desarrollaron en el
hocico las protuberancias óseas que los caracterizan. Se han
encontrado esqueletos de Brontops con heridas
a medio cicatrizar en las costillas, lo cual apoya la teoría
de que los machos utilizaban estas prominencias craneanas en
las luchas por el dominio. Las heridas sugieren que el
animal había recibido en los flancos un golpe muy fuerte que
le propinara un rival, ya que allí no había en esa época
ningún otro animal que fuera capaz de infligir un dańo
semejante. El movimiento de la caja torácica durante la
respiración habrá impedido que los huesos rotos se uniesen
de forma adecuada. Uno de los famosos esqueletos de
Brontops se encontró en unas rocas que se habían
formado en una ciénaga. Es evidente que el animal vivía en
un bosque Cenagoso y que murió al quedar atrapado en el
lodo.
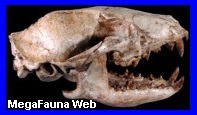
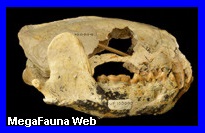
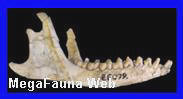
América del Norte. Es un género extinto de
mamíferos euterios de la familia Leptictidae. Ha vivido en
Norteamérica entre los período Oligoceno y Eoceno superior
Chattiano de Estados Unidos. Estaba emparentado con el Leptictidium, un género muy común. Algunas de
estas especies fueron estudiadas y descritas por Joseph
Leidy en la segunda mitad del siglo XIX. Otros análisis más
recientes emparentan este género con las musarańas
elefantes, más que con los insectívoros. Se conocen fósiles
en los estados del centro-norte de Estados Unidos, de los
periodos Oligoceno y el Cretácico tardío, de las especies Leptictis acutidens, Leptictis haydeni,
Leptictis leidy, Leptictis dakotensisSobre
todo, piezas dentales y diversas partes del cráneo.
Andrewsornis abbotti.Patterson,
1941.
Cráneo recreado a partir de los restos encontrados en la
Quebrada Fiera, inmediaciones de Malargüe.
Museo de la ciudad de Mendoza.
Aspecto Andrewsornis de la
gigantesca ave hallada en el Oligoceno de la Provincia de
Mendoza. (*)
Patagonia Argentina.
Vivió en Oligoceno medio y superior de Argentina. Tenia 1,70 metros de
altura. Reconocido como un
Phorusrhacidae es un clado extinto de aves cariamiformes conocidas
también como aves del terror, porque sus especies más grandes eran
depredadores en las diversas regiones que habitaron. Eran aves de
gran tamańo, carnívoras y no voladoras; fueron los depredadores
dominantes en América del Sur durante el Cenozoico, entre 62 y 2,5
millones de ańos. Sus
alas habían evolucionado para utilizarlas como brazos para voltear
una presa en movimiento. Eran corredores rápidos. Sus parientes más
cercanos hoy en día son las chuńas de la familia Cariamidae. Restos
de esta ave fueron exhumados en el Oligoceno de Quebrada Fiera,
inmediaciones de Malargüe,
provincia de Mendoza.
Physornis fortis.
Ameghino, 1895.
Imagen ilustrativa de Paraphysornis brasiliensis,
emparentado directamente con Physornis fortis del
Oligoceno de Argentina. Esqueleto montado en el MEF.
Aspecto en vida
de Physornis fortis. (*)
Patagonia Argentina.
Fue una especie extinta de aves gigantes no
voladoras depredadores de la familia Phorusrhacidae o "aves del
terror", dentro de la subfamilia Brontornithinae, que vivían en
Argentina. Vivió entre los 28 a 23 millones de ańos, durante el
Oligoceno. Sus restos fósiles proceden de la provincia de Santa
Cruz.
Restos fósiles del pingüino
Anthropornis nordenskjoldihallados en Tierra
del Fuego. (*)
Humero de
Anthropornis sp. Museo de La Plata.
Aspecto del primitivo pingüinoAnthropornis nordenskjoldi del Oligoceno del sur Patagonico.
(*)
Patagonia Argentina.
Los pingüinos actuales
constituyen un grupo muy particular de aves no voladoras, adaptadas
para la natación y buceo, preferentemente en aguas frías y
templadas. Su distribución geográfica se restringe al hemisferio sur
en varias regiones entre la Antártida y las islas Galápagos. De
manera semejante a su distribución actual, los hallazgos de
pingüinos fósiles también están restringidos al hemisferio sur, y en
particular el conocimiento paleontológico de este grupo de aves está
basado en material fósil registrado en Nueva Zelanda, Australia,
Sudáfrica, Antártida y Argentina. Los pingüinos fósiles más
antiguos, aproximadamente entre 50 y 40 millones de ańos, se
conocían para Nueva Zelanda y Antártida.
Algunos de ellos fueron verdaderos gigantes y alcanzaron una talla
entre 1,6 y 1,80 m, como el Anthropornis nordenskjoeldi,
una especie fósil hallada en la isla Marambio (Seymour), Antártida,
por la célebre expedición de Otto Nordenskjöld en 1903, en la cual
también participó el argentino José M. Sobral. En la actualidad la
isla Marambio, situada en el extremo norte de la península
Antártica, se encuentra separada del continente americano por el
pasaje de Drake, y aproximadamente a unos 1.200 km de distancia de
Tierra del Fuego. Sin embargo, hace unos 40 millones de ańos, edad
del material fósil fueguino, el pasaje de Drake no existía y la
península Antártica estaba físicamente conectada al extremo austral
de América del Sur. El hallazgo en Tierra del Fuego de pingüinos
fósiles de edad semejante a la del material de la isla Marambio en
Antártida es consistente con esta antigua geografía y apoya otras
evidencias climáticas del pasado, por ejemplares el marcado
incremento en la abundancia de polen de Nothofagus y
las seńales isotópicas en conchillas marinas, que indican el
comienzo de una larga y continua tendencia de enfriamiento de las
aguas oceánicas australes hacia los 40 millones de ańos. A pesar
de la asociación popular de los pingüinos con las frías regiones
polares, existen especies de estos pájaros que viven también más
cerca del ecuador. Los más primitivos, por tanto, debieron haberse
desarrollado en climas más cálidos, para adaptarse lentamente a sus
hábitats cuando éstos se helaron.
Parastrapotherium martiale.
Ameghino 1901.
Rama
mandibular y pieza tipo deParastrapotherium,
colectada por Ameghino y exhibida en el MACN.
Reconstrucción del posible
aspecto (*)
Patagonia Argentina.
Es uno de
los elementos faunístico más característicos de la Edad Mamífero
Deseadense (Oligoceno tardío) de Patagonia.
Parastrapotheriumreveló que este género sólo presenta un
premolar inferior más que Astrapotherium, el cual es muy
semejante a este ultimo. Los astrapoterios, constituyen un orden
reducido pero bien representado de herbívoros de cuerpo bajo, que
sobrevivieron desde fines del Paleoceno hasta mediados del Mioceno.
Algunos de ellos eran tan grandes como los rinocerontes y tenían
colmillos y una trompa pequeńa. Era herbívoro, alimentándose
principalmente de raíces, las cuales encontraba a orillas de los paleopantanos donde pasaban la mayor
parte de su vida. Poseía un par de caninos bien desarrollados y una
pequeńa trompa. Parastrapotherium holmbergi y
Parastrapotherium ephebicum.
Pyrotherium
romeroi.
Ameghino, 1889.
PyrotheriumPyrotheriumPyrotherium
PyrotheriumPyrotheriumPyrotherium
Cráneo de Pyrotherium
en el MEF.
Mandíbula. Foto
Conicet.
Reconstrucción
de Pyrotherium romeroi. (*)
Patagonia Argentina.
Tenía unos 3 metros
de largo y pesaría tres tonelada y media. Su nombre significa
"Bestia de fuego" ya que los primeros restos hallados por Florentino
Ameghino en 1889, provenían de depósitos sedimentarios de origen
volcánicos. Algunos mamíferos fósiles de Sudamérica eran extrańos,
pues, evolucionaron aislados del resto de los continentes.
Pyrotherium se veía como un elefante primitivo en algunos
aspectos, pero poseía características propias muy llamativas. Tenía
patas muy fuertes para soportar el enorme peso de su cuerpo. Su
cabeza estaba dotada de una trompa y sus mandíbulas estaban
acompańadas por seis colmillos, no dos como los elefantes modernos o
como algunas formas fósiles. Eran cortos y en forma de cincel, los
cuales eran utilizados para excavar la tierra en busca de raíces y
bulbos. Los
esqueletos se conocen imperfectamente, las especies más comunes
halladas muestran un cuerpo alargado y bajo. Algunas características
recuerdan al grupo de grandes herbívoros primitivos, los
xenungulados del Paleoceno, con los que se supone que están
emparentados. Sin embargo, el estudio completo del tarso de
Pyrotheriumno apoya esta relación. En un estudio, no se
observaron las características más derivadas del mismo en otros
mamíferos examinados exceptuando al Arsinoitheriumdel
Paleógeno de África.
Sus restos, hallados en Sudamérica (Brasil, Argentina
y Perú), datan del Oligoceno.
Nesodon conspurcatus.Ameghino, 1887.
Vista inferior del cráneo
in-situ. (*)
Cráneo
completo de Nesodon en el MEF.
Aspecto del extinto
Nesodon. (*)
Patagonia Argentina.
Es un género extinto de mamíferos notoungulados de la
familia Toxodontidae que vivieron en el Oligoceno - Mioceno medio.
El aislamiento de América del Sur permitió la
evolución separada de estos grupos de mamíferos, muchos de los
cuales se diversificaron en nichos ecológicos que, en otras partes
del mundo, ocuparon otros grupos. El Nesodon se
parecía a los rinocerontes y los hipopótamos. La relación entre
ellos se confirma tanto por la distribución de las cúspides de los
dientes como por los huesos del oído, que son exclusivos de este
orden de mamíferos. Nesodon se tenía el cuerpo robusto
y grande.
Nesodon,
en el continente sudamericano. Alcanzaba un tamańo relativamente
grande, pesando más de 600 kilogramos y alcanzaba 1.5 metros de
altura a los hombros. El aspecto de estos animales era
impresionante. Su cuerpo era parecido al de un bisonte, con la
espalda arqueada y dotada de grupa. Las pezuńas eran pequeńas y la
cabeza de gran tamańo, casi desproporcionada con el resto del
cuerpo.Nesondon medía aproximadamente 2 metros,
siendo un herbívoro que probablemente viviera en las praderas
sudamericanas. Se
alimentaba de vegetales en zonas áridas. Es una de las especies más
características del Oligoceno de Patagonia Argentina.
Trachytherus spegazzinianus.
Ameghino, 1889.
Cráneo de un
Trachytherus s.p (ilustrativo) del Museo de Historia
Natural de Lima. Oligoceno de Perú.
El técnico
Mariano Magnussen Saffer, junto a restos de
Trachytherus spegazzinianus,
recreados a partir de un cráneo del oligoceno de Patagonia.
Museo Municipal de Miramar.
Aspecto de
Trachytherus s.p. (*)
Patagonia Argentina.El
género Trachytherus tiene un valor crono y
bioestratigráfico muy importante. Se trata de otro genero de los
ungulados nativos sudamericanos o Notoungulata, es decir herbívoros
con pezuńas nativos (a diferencia del ganado actual que también
tienen sus pezuńas). El notoungulado mesoterio Trachytherussegún todos los indicios era un animal con pezuńas muy diferente.
Tenía dientes de crecimiento permanente parecidos a los que
presentan los roedores. Adicionalmente Su distribución parece
circunscribirse solamente al Oligoceno tardío y su distribución
estratigráfica se restringe a la parte superior del «miembro Puesto
Almendra» de la «formación Sarmiento» en la cuenca del golfo de San
Jorge, provincia del Chubut, y a los «estratos de Salla», en la
cuenca de Salla-Luribay, Bolivia. En Patagonia, Pyrotherium
romeroi tiene la misma distribución estratigráfica que la de
Trachytherus spegazzinianus. Se ha recomendado también
a esta especie para caracterizar la fauna del Oligoceno asignada a
la «edad deseadense» en la Patagonia y Bolivia, al contar con un
completo registro estratigráfico. Su paracrón comprende desde
27,6 millones de ańos en Pico Truncado, Argentina, hasta
24,8 millones de ańos en Salla, Bolivia. Era un animal de mediado
tamańo. Usaban sonidos para comunicarse entre ellos, posiblemente
para emparejarse y advertir del peligro. Eran primariamente
herbívoros y se desplazaban según su hábitat, en busca de hojas,
raíces, flores, cortezas, frutos, etc.; también comían insectos y
pequeńos animales. A partir de una copia del cráneo descripto por
Ameghino, el técnico en paleontología Mariano Magnussen Saffer,
recreo parte del esqueleto para una exhibición educativa en el Museo
Municipal Punta Hermengo de Miramar.
Proborhyaena
gigantea.
Ameghino, 1897.
proborhyaena proborhyaena
proborhyaena proborhyae
Esqueleto de borienido, en el MEF.
Cráneo exhibido
in-situ en el parque paleontológico Bryn Gwyn. (*)
Reconstrucción en vida. Tomado
de Prehistoric Fauna.
Patagonia Argentina.
Era parecido a un oso, con las patas planas. Fue un marsupial carnívoro
del orden Sparassodonta
del tamańo de un lobo, de 1,5 a 2 metros de longitud. Como
muchos metaterios, probablemente las hembras poseían una bolsa para
llevar a las crías en desarrollo. Los borhiénidos tenían fuertes y
potentes mandíbulas, capaces de triturar huesos.
Eran depredadores
muy importantes durante el Oligoceno de la patagonia Argentina, los
cuales se alimentaba de mamíferos notoungulados y armadillos. Las patas cortas
seńalan que no era un animal corredor, pero seguro que asechaba a
sus victimas en los antiguos bosques que existían hace 30 millones
de ańos en los actuales desiertos de la provincia de Chubut.
Originalmente, la familia Borhyaenidae fue uno de los grupos más
extensos entre los esparasodontes, incluyendo a todas las especies
que no fueron incluidas originalmente en Thylacosmilidae. Sin
embargo, en ańos recientes, otros grupos de esparasodontes han sido
elevados al rango de familia y esto junto al descubrimiento de que
los borhiénidos están más cercanamente relacionados con los
proborhiénidos y los tilacosmílidos que a otros esparasodontes, la
familia ha sido reducida a seis especies en tres géneros
Cráneo de Tremacebus, un
primate primitivo procedente del Oligoceno. (*)
Cráneo de Tremacebus harringtoni. (*)
Aspecto posible del primate
de la desaparecida selva patagonica. (*)
Patagonia Argentina.
Fue un primitivo primate (monos) aloctono, cuyos ingreso al
continente fue hace 38 millones de ańos según la evidencia fósil
reunida hasta el momento. A diferencia de los monos del Viejo Mundo,
los monos del Nuevo Mundo no parecen haberse dado a la vida en las
praderas y siguieron siendo animales de selva. Una gran diferencia
entre ambos, es que los primeros no poseen cola, mientras que los
sudamericanos poseen unas vértebras caudales extensas y adaptadas
como un quinto miembro para poder trepar. Tremacebus
harringtoni,
es un representante del periodo Oligoceno tardío (23,5 millones de
ańos) en la Patagonia Argentina, y se parecía a un mono Buho
moderno. Otros pequeńos simios del territorio argentino son
Homunculus patagonicus y
Killikaike blakei,
entre otros. Los primates mas primitivos por ahora en América del
sur es Branisella, del oligoceno temprano (33,7 millones de ańos) de
Bolivia. Mientras que dos parientes del actual mono arańa, el
Protopithecus y el Caipora, vivieron en
Brasil durante el Pleistoceno. Era omnívoro y tenia una variada
dieta. Este género fue descrito
originalmente por el paleontólogo Hershkovitz en el ańo 1974. La
única especie en que está basado fue descrita originalmente por el
paleontólogo Carlos Rusconi en el ańo 1933. Se incluye en la familia
de los aótidos (Aotidae), exclusiva del Neotrópico, que comprende un
solo género viviente, con un total de 11 especies, siendo los únicos
monos nocturnos. Carlos Rusconi recibió la muestra en el ańo 1932
enviada por Harrington Thomas, quien lo recogiera junto con otros
restos fósiles de mamíferos de la «edad Colhuehuapense», a unos 12 km
al suroeste del cerro Sacanana, en el centro-norte de la provincia
del Chubut, en el centro de la Patagonia argentina.
Pseudhapalops
observationis.Ameghino, 1891.
Esqueleto casi completo de Pseudhapalops
s.p, exhibido en el Museo de La Plata.
Oligoceno-Mioceno temprano de Santa Cruz.
Copia de cráneo y mandíbula de Pseudhapalops
s.p, en el Museo Municipal Punta
Hermengo de Miramar. Oligoceno-Mioceno temprano de Santa
Cruz.
Aspecto en vida
de Pseudhapalops
s.p, cuando la Patagonia era una selva tropical. (*)
Patagonia Argentina.
A finales del Oligoceno, hace 24 millones de ańos, los Xenarthros
estaban bien representados y adaptados en América del sur, el cual,
en el Mioceno se encontraba geográficamente bien distribuido por la
actual patagonia Argentina. Pseudhapalops
era de tamańo chico, media hasta un
metro de largo, a diferencia de sus gigantescos parientes del
Pleistoceno que superaban los cuatro metros de altura. Tenia garras
muy desarrolladas y curvas (una característica común entre los mamíferos edentados) seguro debe haberse visto desplazarse sobre sus nudillos
anteriores. Es muy probable que pasara la mayor parte de su tiempo
en las copas de los árboles, sujetándose de sus garras y alimentándose
de frutos suculentos en los tupidos bosques sub tropicales del
Oligoceno. Como todos los edentados, Pseudhapalops tenia pocos dientes y
sin esmalte. Sus mandíbulas solo tenían cuatro pares de dientes. Si
bien hoy en día la Patagonia Argentina es un gran desierto, durante
el Oligoceno y Mioceno estaba constituida por grandes y extensas
selvas, evidenciado por la diversidad faunistica y botánica
recuperada. Otras especies relacionadas; Eucholoeops ingens
y Pelecyodon s.p.
Patagonia Argentina.Fue un "Dasipodido" (peludos y mulitas). Tenia una coraza
formada por placas óseas, las cuales avían evolucionado de placas
térmicas, formando un caparazón recubierto de cornea y con bandas
móviles, lo que le brindaba movimientos rápidos. Peltephilus, poseía
en en el hocico un par de cuernos largos, que en vida se encontraban
recubiertos por piel y tejido corneo, lo que facilitaba su
utilización en momentos de defenderse de sus depredadores. Algunos
científicos sostienen que este animalito que tenia unos 60 centímetros
de largo, tal vez poseía otro par de pequeńos cuernos córneos en
el hocico. Con respecto a otros "edentados", Peltephilus
tenia unos dientes gruesos y largos, muy parecidos a caninos, lo que
podría indicar que era un animal carnívoro o carrońero.
Clypeotherium magnum.
ScillatoYané, 1977.
Coraza de Clypeotherium magnum. del Oligoceno de Chubut,
Argentina. Museo de La Plata.
Posible aspecto de Clypeotherium magnum.
Patagonia Argentina.
Este enorme armadillo comparable con los que vivieron
durante el Pleistoceno, pertenece a la familia extinta
Glyptodontidaede mamíferos placentarios del orden Cingulata,
emparentados con los actuales armadillos extintos y actuales.
Originarios de Sudamérica, se diferenciaron en el Eoceno Superior.
Tenían un cráneo y un caparazón formado por muchas piezas pequeńas
fusionadas y, en ocasiones, una cola cubierta. Muestran una notable
convergencia con las tortugas y algunos dinosaurios (anquilosauros).
Tenían una adaptación a la dieta herbívora y conversión de sus
bandas acorazadas del dorso en una suerte de único caparazón
defensivo. Perteneció a la subfamilia de los Glyptatelinae,
gliptodontes primitivos conocidos en el Oligoceno de Argentina, que
aparentemente pudieron sobrevivir de manera relictual en el norte de
Sudamérica en el Neógeno. Su coraza tiene un grosor de 4
centímetros, y por el margen de esta, estaba constituidas por placas
puntiagudas.
Argyrocetus patagonicus.
Lydekker, 1894.
Cráneo y mandíbula del Oligoceno tardío - Mioceno temprano
de Chubut. Museo de la Plata.
Aspecto de un integrante de la familia
Eurhinodelphinidae (*).
Patagonia Argentina.
Los delfines
de la familia extinta Eurhinodelphinidae son odontocetos que
vivieron del Oligoceno hasta el Mioceno. Argyrocetus patagonicus, es el miembro mas
primitivo de esta familia de cetáceos. Los miembros de
familia poseían una prolongación de la mandíbula superior con
apariencia similar a un pez espada. De alimentación carnívora y
oportunista. La historia evolutiva de los cetáceos estuvo
marcada por al menos tres radiaciones adaptativas. La segunda
radiación (Oligoceno-Mioceno Temprano) marcó la aparición de los
Neoceti (Odontoceti y Mysticeti), y estuvo principalmente
influenciada por la apertura del Océano Austral y cambios en el
ecosistema oceánico y su productividad. Algunos de estos
materiales mas modernos, provienen del Oligoceno Tardio - Mioceno Temprano en la
Formación Gaiman y con una antigüedad de 20 millones de ańos
antes del presente.
Carcharodon megalodon.
Agassiz, 1843.
Diente de
Carcharodon megalodon. (*)
Mandíbula de
Megalodon recreada a tamańo natural en el Museo
Paleontológico de Bariloche.
Reconstrucción en vivo (*)
Patagonia Argentina.
Se trata de un enorme tiburón prehistórico que habito en los
antiguos mares del territorio Argentina y distintas partes del
mundo. Poseían un esqueleto cartilaginoso, el cual rara vez se
preserva, pudiendo agregar que los hallazgos de este tipo de
estructura se pueden contar con los dedos de las manos. La única
parte del esqueleto que podemos hallar perfectamente preservado, son
los dientes con esmalte, los cuales varían desde 2 a 15 centímetros.
Su raro esqueleto se distinguen claramente del gran número de
especies de peces teleósteos (óseos) por su esqueleto
cartilaginoso. Los tiburones son peces versátiles y tienen sentidos
muy agudos; muchas de sus especies son capaces de cazar y devorar a
casi cualquier otro animal marino, tanto en mares profundos como en
mares poco profundos. Estos dos rasgos explican su larga historia
evolutiva; muchas de las especies de tiburones de nuestros días son
muy similares a especies que nadaban en los mares del cretácico,
hace más de 100 millones de ańos. Su alimentación estaba
constituida por grandes reptiles marinos de la época, peces y mamíferos
marinos primitivos. Un sitio muy conocido en nuestro país donde se
han hallado restos dentarios de la especie, es Cerro Gualicho en la
Provincia de Rió Negro.
Archaeogeryon peruviensis, Oligoceno superior y Mioceno de
la Provincia de Santa
Cruz, Argentina. (*) El segundo ejemplar
Archeogeryon fuegianus
se exhibe en el
Museo Paleontológico Egidio Feruglio de Trelew (MEF):
Patagonia Argentina.
Se trata una especie de cangrejo marino muy abundante en el
registro fosilífero, aunque generalmente solo se hallan pequeńos
fragmentos, y rara vez formas completas. Estos vivían en los
antiguos mares que se hallaban sobre los actuales sedimentos de Patagonia. Poseen un cuerpo similar a las especies vivientes,
formado por reducción del abdomen y el ensanchamiento y aumento de
tamańo de la parte anterior (delantera) del cuerpo. El abdomen
sirve como bolsa de cría para los huevos, que en el hembra es mas
notorio por su tamańo. El cuerpo está más o menos cubierto por un
caparazón quitinoso, con una cubierta cerúlea. El cuerpo
segmentado del cangrejo tiene varios pares de apéndices, de los
cuales cinco suelen servir para la locomoción y dos hacen las veces
de antenas sensoriales. Las patas delanteras estaban equipadas con
pinzas que le servían para alimentarse, defenderse y realizar
exhibiciones rituales de apareamiento. Los cangrejos son capaces de
tolerar cambios en su medio ambiente, los que les permite prosperar
en hábitats muy hostiles, por ello se lo considera junto a los artrópodos
como formas exitosas y primitivas. Sus hábitos alimentarios
son variados. Algunos fósiles de
Archeogeryon fuegianusfueron hallados muy
completos recientemente en los afloramientos geológicos del
Oligoceno de Península de Valdes en la Provincia de Chubut, como así
también en sedimentos de Caleta Olivia en la Provincia de Santa
Cruz.
Egipto, Malí y Senegal.
Uno de los fósiles más antiguos de mamíferos debe ser el Arsinoitherium, un extrańo animal parecido al
rinoceronte. Se le conoce por medio de cráneos y esqueletos
bien conservados de los mismos depósitos del
Moeritherium, en Egipto, y algunos otros más
deteriorados del Medio Oriente. Su característica más obvia
son los dos grandes "cuernos" que se levantan sobre el
hocico. Se funden en la base y son huecos. En los machos
eran puntiagudos y en las hembras y jóvenes más pequeńos y
romos. En los animales vivos, los cuernos estaban cubiertos
de piel, lo que se deduce de la presencia de vasos
sanguíneos en la parte exterior del hueso, así que deben
haberse visto más como los cuernos de una jirafa que como
los de un rinoceronte. El Arsinoitherium tenía
dientes altos, probablemente para triturar plantas duras.
Otras especies:Arsinoitherium andrewsii,
Arsinoitherium giganteumy
Arsinoitherium zitteli.
África (Egipto). Tenia
2.5 m de altura Phiomia evolucionó al mismo tiempo que su
primo lejano, más pequeńo, Moeritherium,
aunque es poco probable que compitieran por el mismo tipo de
alimentos. Lo más factible es que Phiomia
ramoneara en los bosques, mientras su contemporáneo se
bańaba en las ciénagas. Su nombre quiere decir «criatura de
la provincia del lago», por referencia a la zona de Fayún,
en Egipto, donde se encontraron sus restos. Tanto la
mandíbula superior como la inferior eran largas, y los
colmillos aplanados del maxilar inferior formaban una
prolongación acucharada que se utilizaba para juntar los
alimentos. Es probable que el labio superior se extendiera
para formar una trompa primitiva que funcionara junto con
este extrańo aparato de la mandíbula superior. Los colmillos
más cortos de la mandíbula superior tal vez se utilizaran
como armas defensivas.
Senegal y
Malí. Durante mucho tiempo se consideró que el
Moeritherium era el elefante más antiguo que se
conocía, pero en 1984 se reportaron restos aún más viejos en
Argelia. Están mucho menos completos que los ejemplares de
Moeritherium, algunos de los cuales provienen
de los mismos yacimientos de Egipto de donde salieron
esqueletos de Arsinoitherium. El
Moeritherium probablemente se veía como un
hipopótamo pigmeo y carecía de colmillos y trompa. Los
molares y otras características del cráneo y del esqueleto
muestran que era un elefante primitivo, pero probablemente
una línea lateral de la que siguieron las formas modernas.
El Moeritherium tenía largos dientes frontales
que sobresalían, tanto en la mandíbula inferior como en la
superior, parecidos a los colmillos dobles que se observan
en los gomfodontes, animales de mayor tamańo que surgieron
después. Los fósiles de Moeritherium provienen
de rocas de la boca de un río cercana al mar, por lo que
debe haber vivido buena parte del tiempo en el agua,
arrancando plantas acuáticas con sus dientes.
Asia
(China), Europa (Francia) y América del Norte (Estados
Unidos). Tenia 75 cm de longitud. Este cazador
arborícela tenía los ojos grandes, tal vez para poder cazar
de noche, y una cola larga que le servía para mantener el
equilibrio. Era semejante al basarisco (Bassctriscus
sumichrasti) actual, y es posible que fuese su
antepasado directo. Tal vez llevaba la misma forma de vida,
subiendo y bajando por los troncos de los árboles, saltando
de un árbol a otro y corriendo por las ramas. Como en todos
los prociónidos, las cúspides de los dientes eran romas, y
los molares eran cuadrados, si se examinaban en sentido
transversal, lo cual indica que es probable que tuviera una
alimentación omnívora, devorando los huevos de las aves,
insectos y plantas, aparte de cazar mamíferos pequeńos y
aves.
Europa (Francia y Alemania) y
América del Norte (Nebraska). Tenia 2 m de
longitud Amphicyon era un «perro oso» típico.
Es probable que se pareciera a un oso de gran tamańo, con
los dientes fuertes y afilados de un lobo. Tenía el cuello
grueso, patas poderosas y una cola pesada. Es posible que
viviera como los osos pardos o grises modernos,
alimentándose de una gran variedad de plantas y animales, y
matando a sus presas con fuertes golpes de las patas
delanteras. Amphicyon fue sin duda un
adversario temible para cualquier otra criatura que viviera
en las planicies del hemisferio norte a mediados del
Terciario, hace unos 30 millones de ańos. Una especie fósil,
Amphicyon giganteus, que vivió en Europa
durante el Mioceno, era del tamańo de un tigre actual.
Hoplophoneus
(Eusmilus) sicarius.
Sinclair and Jepsen,
1927.
Europa (Francia) y América del Norte (Colorado, Nebraska,
Dakota del Norte y del Sur y Wyoming). Tenia
2,5 metros de longitud. Este felino del tamańo de un
leopardo tenía el cuerpo bastante largo y las patas bastante
cortas si lo comparamos con los felinos modernos. Apareció
por primera vez en Europa hacia el final del Eoceno, hace
unos 40 millones de ańos, y se difundió hacia el este, a
América del Norte, a través del istmo de Bering, durante el
Oligoceno. Eusmilus era un miembro
característico del grupo de los félidos de dientes de sable.
El par de caninos superiores se había alargado para formar
unos «sables» afilados y bien desarrollados. Los caninos
inferiores eran insignificantes y había perdido muchos de
los otros dientes. Eusmilus sólo conservaba 26
dientes en sus mandíbulas, en comparación con el máximo de
44 que aparecen en algunos carnívoros. La articulación de la
mandíbula estaba modificada para abrirse hasta formar un
ángulo de 90°, gracias a lo cual los dientes de sable podían
realizar su trabajo. Las mandíbulas inferiores tenían unas
cubiertas óseas que protegían a los «sables» cuando se
cerraba la boca. En esto se parecían a Thylacosmilus,
el diente de sable marsupial de Argentina, aunque estos
mamíferos no estaban relacionados entre sí. La similitud es
un ejemplo de evolución convergente. Eusmilus y otros falsos felinos de dientes de
sable habitaban los mismos lugares del mundo, al mismo
tiempo, y existen pruebas fósiles de que sus caminos se
cruzaron. En América del Norte se ha encontrado un cráneo de
Nimravus con un agujero en la región frontal
que corresponde exactamente a las dimensiones del diente de
sable de un Eusmilus. Sin embargo, la herida
no fue fatal, ya que Nimravus sobrevivió a la
pelea durante el tiempo suficiente para que la herida
cicatrizara.
América del Norte (Nebraska y Dakota del Sur).
Tenia 3 metros de longitud. Los enrelodontos alcanzaron su
mayor tamańo con el omnívoro Dinohyus, de
América del Norte. Este animal era muy parecido a.
Archaeotherium, pero tenía un tamańo similar al de
un toro. Si bien tenía las mismas proporciones corporales
que el cerdo, se supone que la cara debía de ser bastante
diferente; por ejemplo, la nariz no era plana y las fosas
nasales se abrían a- os lados del hocico y no en la parte
anterior. Como ,Archaeotherium, Dinohyus
se alimentaba cerca del suelo, compensando con el
hocico largo la escasa longitud del cuello.
Europa y Asia. Los
rinocerontes hiracodontos sin cuernos, de los que existen
alrededor de una docena de géneros, fueron la familia más
antigua y más primitiva del grupo. Es probable que
evolucionaran a partir de un tapiroide próximo a
Hyrachyus. Las muelas, largas y eficaces, eran
similares a las de los tapires, pero los incisivo y los
caninos estaban modificados de distintas maneras. Los
hiracodontos primitivos tenían una estructura parecida a la
de los caballos y las extremidades delgadas y alargadas. Los
miembros posteriores de esta familia desarrollaron una forma
más robusta. Todos sabemos que el mayor animal que camina
hoy día sobre tierra firme es el elefante africano,
Loxodonta africana, protagonista de tantos documentales que
cualquier dato que pudiera yo dar aquí resultaría
completamente redundante. Pero, en el pasado, caminaron
seres mucho mayores sobre la Tierra. El honor de ser el
mayor mamífero terrestre es disputado entre dos herbívoros
estrechamente relacionados, los rinocerontes gigantes del
Mioceno y Oligoceno, como el Indricotherium y
el Paraceratherium, este ultimo, con una
altura máxima de unos 6 m hasta los hombros, una longitud de
unos 9 m y un peso de 18 o 20 Toneladas. En comparación, los
mayores machos de elefante africano pesan hasta 6 Tm, con
una altura de hasta 4 m. Estos antecesores de los
rinocerontes y los tapires, relacionados también con los
actuales caballos, llevaban posiblemente una vida muy
parecida a la de los elefantes de hoy en día, con la
salvedad obvia de que podían comer hojas de árboles más
altos. Se encuentra ligeramente relacionado con el genero Forstercooperia.
América del Norte y Europa.
Es un
género
extinto de
mamíferos creodontos de la
familia
Oxyaenidae que vivió desde el
Paleoceno Superior hasta el
Eoceno Inferior en
América del Norte y
Europa.
Se cree que era un animal omnívoro, pero que dieta se basaba
en carne. Eran animales similares a gatos con locomoción
plantígrada, al contrario de los
carnívoros actuales (a excepción de los
osos y
mapaches) que son
digitigrados. Se caracterizaban por un cráneo corto y
ancho, mandíbulas fuertes y dientes diseńados para triturar
en lugar de cortar, a diferencia de los
hienodóntidos. Eran carnívoros especializados que se
alimentaban de aves, mamíferos pequeńos, huevos e insectos;
la evidencia fósil sugiere que eran capaces de trepar a los
árboles.
Asia (Mongolia). Los anagalidios eran mamíferos
parecidos a los conejos, escarbadores, que se conocen desde
el comienzo del Terciario, en el este de Asia. En una época
se creyó que estaban relacionados con las musarańas
elefantes, pero hoy se considera que tienen más en común con
los roedores y los conejos. Es posible que Anagale se pareciera a los conejos actuales, pero tenía una cola
larga y, probablemente, orejas cortas. Además, corría en vez
de saltar como hacen los conejos. Esto se deduce de las
proporciones de los huesos de sus patas traseras. Las patas
traseras de Anagale eran un poco más largas
que las delanteras, y los pies estaban equipados con unas
garras que parecían espadas. Es probable que Anagale
revisara el suelo en busca de escarabajos, orugas, gusanos y
otros insectos. El hecho de que, en muchos de los
especimenes que se han encontrado, los dientes estuvieran
muy gastados, sugiere que tal vez se comiera también la
tierra, lo cual tal vez sea el método más sencillo para
extraer de ella los alimentos.
Asia (Mongolia). Tenia 2,5 metros de altura.
La cabeza de Embolotherium presenta las formas
grotescas típicas que desarrollaron los últimos brontoterios.
Desde la parte posterior del cráneo se prolongaba hacia
adelante formando un hueco profundo y después se elevaba en
un «cuerno» único situado sobre la nariz. Los ojos estaban
situados en la parte anterior, justo detrás de las fosas
nasales, y en la base del cuerno. El cráneo angosto dejaba
poco sitio para el cerebro; como en otros brontoterios
enormes, el cerebro no era más grande que el puńo humano. La
aparición de Embolotherium en el desierto de
Gobi, en Asia, indica lo difundidos y prósperos que fueron
los brontoterios en su época de mayor auge.
Asia (Pakistán y China).
Tenia 8 metros de longitud. Parece imposible que un animal
tan pequeńo, ligero y ágil como Hyracodon
pudiera convertirse en el mamífero terrestre más grande que
se conoce, aunque todas las pruebas apuntan en esa
dirección. Indricotherium, también conocido
con el nombre de Baluchitherium, por el estado
pakistaní en donde se encontraron los ejemplares más
importantes, era un animal inmenso. Su peso se estima en 30
toneladas, el doble que el mamut más grande que se conoce y
más de cuatro veces el peso del elefante moderno. Sólo el
cráneo medía 1,3 m de largo, pero su tamańo era pequeńo en
relación con el cuerpo en general. Las vértebras dorsales y
cervicales estaban llenas de agujeros y puntales, como las
de los mayores dinosaurios, para aligerar el peso
conservando la fuerza. Las patas eran como las de los
elefantes, pero parece que el peso del cuerpo lo soportaban
apenas tres dedos, como suele ocurrir en los rinocerontes.
Tampoco tenía cuernos; en realidad, los huesos nasales eran
bastante endebles. Los dientes anteriores de los
rinocerontes fósiles varían considerablemente, pero los de
Indricotherium eran, sin ninguna duda, muy
extrańos. Sólo tenía dos dientes frontales en la parte
superior y otros dos en la parte inferior; el par superior
estaba dirigido hacia abajo, como si fueran colmillos,
mientras que el par inferior iba hacia adelante. Como
existen algunas pruebas de que tenía un labio superior
grande y flexible, es probable que esta disposición haya
posibilitado que Indricotherium ramoneara,
como las jirafas, en las copas de los árboles, a más de 8 m
del suelo. Es probable que Indricotherium
viviera en pequeńos grupos familiares, aprovechando los
árboles dispersos que quedaban en las llanuras abiertas y
áridas. Se halló un esqueleto en unas rocas formadas por
barro cenagoso. Resulta fácil imaginar las dificultades que
una criatura tan inmensa habrá encontrado en un pantano.
Nueva Zelanda. Fue un
pingüino gigante vivió hace 27 a 24 millones de ańos, cuando
Nueva Zelanda estaba en su mayor parte bajo el agua y estaba
constituida por afloramientos rocosos aislados, que les
ofrecían a estos animales protección contra los depredadores
y abundancia de alimentos, dijeron los investigadores.
Los
primeros rastros de los pingüinos, llamados 'Kairuku'
–palabra maorí para denominar al nadador que regresa con la
comida– fueron encontrados en un acantilado en Waimate, en
la Isla Sur, en 1977, por el paleontólogo Ewen Fordyce, de
la Universidad de Otago. Con los ańos, Fordyce descubrió
restos más completos e invitó en 2009 al especialista de la
Universidad de Carolina del Norte, de EEUU, Dan Ksepka para
que lo ayudara a reconstruir el animal. Ambos expertos
determinaron que el ave era mucho más grande que el mayor de
los pingüinos modernos, el emperador, que alcanza hasta un
metro, y pesaba hasta 60 kilos, el doble que el emperador.
Kairuku era un ave elegante para los estándares de
pingüinos, con un cuerpo delgado y aletas largas, aunque
tenía piernas y pies cortos y gruesos. Esta ave de gran
tamańo estaba adaptada para poder nadar más y bucear más
profundo que sus pares actuales.
Benton, Michael. (1992).
Dinosaurios y otros animales prehistóricos. Ediciones
Lrousse Argentina S.AI.C.
Bond,
M.; López,; G.M.; Reguero, M.A.; Scillato-Yané, G.J.; Vucetich, M.G.
1998. Los mamíferos de la Formación Fray Bentos (Edad Deseadense,
Oligoceno superior?) de las provincias de Corrientes y Entre Ríos,
Argentina. Asociación Paleontológica Argentina, Publicación
Especial, Vol. 5, p. 41-50.
B. Lander. 1998. Oreodontoidea. In C. M. Janis,
K. M. Scott, and L. L. Jacobs (eds.), Evolution of Tertiary
mammals of North America 402-425
Barry Cox, Colin Harrison, R.J.G.
Savage, Brian Gardiner. 1999. The Simon & Schuster Encyclopedia of
Dinosaurs and Prehistoric Creatures: A Visual Who's Who of
Prehistoric Life.
Evans, S.E.; Hecht, M.K. (1993). «A history of an
extinct reptilian clade, the Choristodera: longevity, Lazarus-Taxa,
and the fossil record». Evolutionary
Biology27:
323-338.
Forster Cooper, C. (1962). Giant rhinoceros from
Loping, Yunnan, and discussion on the taxonomic characters of
Indricotherium grangeri. Vert. Pal. Asiat. 6 (1), pp. 57-71.
Gómez, Raúl O.; Báez, Ana M.; Muzzopappa, Paula
(10 de febrero de 2011). «A new helmeted frog (Anura:
Calyptocephalellidae) from an Eocene subtropical lake in
northwestern Patagonia, Argentina». Journal
of Vertebrate Paleontology31 (1):
50-59.
Magnussen Saffer,
Mariano (2013).
Herbívoros prehistóricos representativos durante el Oligoceno.
Paleo, Revista
Argentina de Paleontología. Boletín Paleontológico. Ańo 11.
89: 10-15.
Magnussen Saffer, Mariano (2013). Los principales representantes
de la paleofauna del Oligoceno de la Republica Argentina. Paleo,
Revista Argentina de Paleontología. Boletín Paleontológico. Ańo
11. 82: 11-17.
Magnussen Saffer, Mariano
(2013). Mamíferos carnívoros representativos durante el
Oligoceno. Paleo, Revista Argentina de Paleontología.
Boletín Paleontológico. Ańo 11. 88: 17-22.
Magnussen Saffer, Mariano (2013).
Herbívoros prehistóricos representativos
durante el Oligoceno. Paleo,
Revista Argentina de Paleontología. Boletín Paleontológico.
Ańo 11. 89: 10-15.
Matsumoto, H. 1922. Revision of
Palćomastodon and Mśritherium. Palćomastodon intermedius, and
Phiomia osborni, new species. American Museum Novitates. Number 51,
November 21.
Matsumoto, H. 1923. A Contribution to
the Knowledge of Mśritherium. Bulletin of the American Museum of
Natural History ; v. 48, article 4. p. 97-140.
M. Mendoza, C. M. Janis y P. Palmqvist (2006). «Estimating
the body mass of extinct ungulates: a study on the use of
multiple regression». Journal
of Zoology.
Rodolfo Salas, José Sánchez, César Chacaltana.
2006. A New Pre-Deseadan Pyrothere (Mammalia) From Northern Peru
And The Wear Facets Of Molariform Teeth Of Pyrotheria. Journal
Of Vertebrate Paleontology,
26(3):760-769
Paolo
Pirasa and Angela D. Buscalionib (2006). «Diplocynodon
muelleri comb. nov., an Oligocene diplocynodontine
alligatoroid from Catalonia (Ebro Basin, Lleida Province, Spain)». Journal
of Vertebrate Paleontology26 (3): 608-620
Soria, M.F. (h). 1981. Los Liptoterna del Colhuehuapense (Oligoceno
Tardío) de la Argentina. Revista del Museo Argentino de Ciencias
Naturales “Bernardino Rivadavia”, Serie Paleontología 3: 1-54.
Soria, M.F. (h). 2001. Los Proterotheriidae (Litopterna, Mammalia),
sistemática, origen y filogenia. Monografías del Museo Argentino de
Ciencias Naturales 1: 167 pp.
Vizcaíno SF, Farińa RA. 1997. Diet and locomotion in
Peltephilus: A new view . Lethaia 30 (1): 79-86
Schlosser Max (1911) "Beiträge zur Kenntnis der
oligozänen Landsäugetiere aus dem Fayum, Ägypten." Beiträge zur
Paläontologie und Geologie Österreich-Ungarns,
24: 1–167.
Shockey, B.J. 1997.
Toxodontia of Salla, Bolivia (Late Oligocene): Taxonomy,
systematics, and functional morphology.
Ph.D. Dissertation (Unpublished),
University of Florida, p. 1-277.
Thorpe, M. R. (1921). «John Day Eporeodons,
with descriptions of new genera and species». American
Journal of Science.
Ver mas
bibliografía utilizada para hacer la presente Pagina; AQUÍ.
Ver bibliografía de divulgación sugerida; AQUÍ.