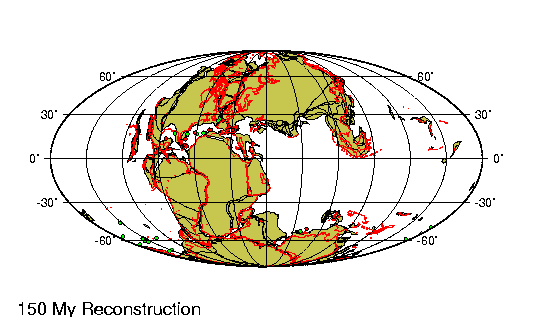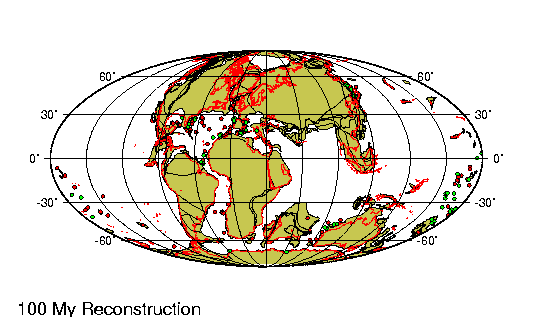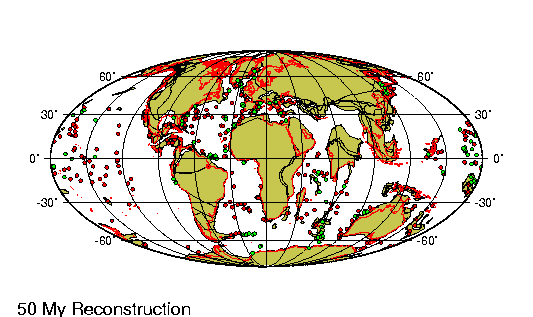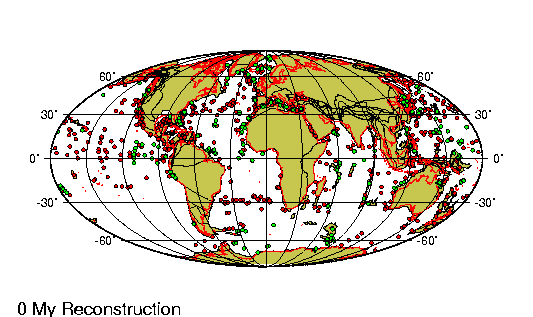
El Holoceno (del griego holos,
todo, y kainos, reciente: la era totalmente reciente),
una división de la
escala temporal geológica, es la última y actual
época geológica del
período Cuaternario. Comprende los últimos 11.784 ańos,
desde el fin de la última
glaciación.
Es un período interglaciar en el que la
temperatura se hizo más suave y la capa de hielo se
derritió, lo que provocó un ascenso en el nivel del mar.
Esto hizo que
Indonesia,
Japón y
Taiwán se separaran de
Asia;
Gran Bretańa, de la
Europa continental y
Nueva Guinea y
Tasmania, de
Australia. Además, produjo la formación del
Estrecho de Bering. La única
especie humana que ha vivido en está época ha sido el
Homo sapiens, que durante estos últimos milenios
desarrolló la
agricultura y la
civilización, ocasionando importantes cambios en el
medio ambiente. La
deriva continental a lo largo de estos casi 12.000 ańos
ha sido de menos de un km, lo que es casi irrelevante. Sin
embargo, el hielo se derritió causando que el nivel
del mar subiera unos 35 m durante esta época y 120 m
desde el último máximo glaciar, hace alrededor de
20.000 ańos. El aumento del nivel no se produjo de
forma uniforme a lo largo de este tiempo, sino que
hubo varios pulsos de rápido deshielo, alternados
por otros de deshielo progresivo.
La mayor parte del aumento del nivel del mar se produjo
antes de 6.000 ańos atrás. El Holoceno comenzó después de que
finalizara el evento de enfriamiento del Dryas Reciente, a partir del cual las temperaturas se hacen más
suaves. El
Óptimo Climático del Holoceno fue un período cálido que se produjo
durante el intervalo comprendido aproximadamente entre 9000 y 5000 ańos
atrás. El clima mundial era entre 0,5-2 °C más cálido de lo que es
actualmente. Sin embargo, el calentamiento probablemente no fue uniforme
en todo el mundo. Este período terminó hace alrededor de 5.500 ańos,
cuando las primeras civilizaciones humanas en Asia y África empezaron a
florecer.
A continuación, comenzó el Neoglacial, en el cual las temperaturas disminuyeron progresivamente
hasta el siglo XX, con la posible excepción del calentamiento del
Óptimo Climático Medieval (siglos X a XIV). El punto culminante del
enfriamiento fue la
Pequeńa Edad de Hielo, con tres máximos: sobre 1650, 1770 y 1850. La vida animal y vegetal no ha
evolucionado mucho durante el corto Holoceno, pero se han
producido importantes cambios en la distribución de plantas
y animales. Un gran número de animales, incluidos los
mamuts,
mastodontes, tigres con dientes de sable como Smilodon y
Homotherium, y los perezosos gigantes
desaparecieron en Norteamérica. En este continente
también se extinguieron animales que sobrevivieron
en otras partes, incluyendo caballos y camellos.
Esta extinción de la megafauna americana
coincide con la llegada del ser humano hace 12000 ańos, al
igual que las extinciones anteriores de grandes marsupiales
en Australia hace 40000 (como el Diprotodon) o las posteriores de aves no voladoras en
Nueva Zelanda hacia el ańo 1300 d.c (como el
moa). Las extinciones de
plantas y animales continúan hoy en día. La tasa observada
de extinción se ha acelerado de manera espectacular en los
últimos 50 ańos. En general, el evento de extinción del
Holoceno se caracteriza por deberse a factores relacionados
con la presencia humana y por producirse en un tiempo
geológico muy corto (decenas de miles de ańos) en
comparación con la mayoría de los otros eventos de
extinción. A veces se la denomina la sexta extinción, pues
anteriormente hubo
cinco grandes eventos de extinción.
América
África
Europa
Asia
Oceanía
Camelops
Raphus
Panthera
Palaeopropithecus
Castoroides
Hippopotamus
Thylacinus
Bison
Cygnus
Harpagornis
Ornimegalonyx
Pinguinus
Aptornis
Pygoscelis
Chelychelynechen
Canis
Mekosuchus
Dusicyon
Daubentonia
Eutatus
Dinornis
Pontoporia
Camelops sulcatus.
Cope 1893.
México, Estados Unidos, Canadá y Alaska. Es un
género
extinto de mamíferos artiodáctilos de la
familia Camelidae, propio del oeste de
Norteamérica, de donde desapareció al final del
pleistoceno hace unos 10 mil ańos. La razón de su
desaparición es pobremente entendida pero fue parte de la
gran extinción de megafauna norteamericana, en la cual
caballos,
mamuts, mastodontes,
megaterios,
Panthera leo atrox, los osos Arctodus,
felinos dientes de sable y un gran número de animales de
grandes dimensiones se extinguieron al fin de la última
glaciación. Camelops hesternus tenía un
tamańo máximo de un poco más de 2.10 m de altura al hombro,
haciéndole ligeramente más alto que el moderno Camello
bactriano. Restos de pastos y otras plantas encontradas en
su dentadura sugiere que éste animal comía cualquier hierba
disponible, de la forma en que lo hacen los camélidos
actuales. Debido a que los tejidos blandos generalmente no
se preservan en los restos, no hay certeza de si Camelops
poseía una joroba como los camellos del Viejo Mundo o una
espalda plana como su pariente suramericano, la llama.
Castoroides ohioensis.
Foster, 1838.
Estados Unidos y
Canadá. Es una
especie
extinta de
roedor de la
familia Castoridae. Es uno de los roedores más grandes que jamás
han existido, que alcanzaban el tamańo de un
oso. Vivió hace aproximadamente 50 000 ańos durante el
Pleistoceno en
América del Norte. Utilizaba sus
dientes para roer vegetación dura como cortezas de
árboles, era buen nadador, pero no construía presas. Otros
tipos de
castor construyen presas para protegerse de los
depredadores pero en este caso el propio tamańo agigantado
lo protegía, han sido encontrados fósiles de Castor gigante
desde
Florida (sur de Estados Unidos) hasta el norte de
Canadá.
Bison priscus.
Bojanus , 1827.
Europa,
Asia, Beringia y Norteamérica. Fue una especie de
bisonte, hoy
extinta, que habitó gran parte de
Europa,
Asia Central, Rusia, Manchuria y
América del Nortedurante el
Pleistoceno. Es probable que
evolucionara originalmente en
Asia, al
igual que muchos
bóvidos
actuales y otros elementos de la fauna
glaciación con los que compartió
hábitat. Se
trataba de animales similares a los actuales
bisontes europeos, pero mucho más robustos y grandes, de cuernos más
largos y adaptados a la vida en espacios abiertos como las
estepas
y los límites de la
tundra.
Sus relaciones con las especies de
bisontes
actuales no están del todo claras, aunque tiende a asumirse que la
especie dio origen al actual bisonte europeo a comienzos del
Holoceno a partir de formas menores como Bison priscus mediator,
que se adaptaron al nuevo medio cálido y boscoso que comenzaba a
sustituir a la tundra-estepa glacial en Europa. Las subespecies gigantes
de Asia y Norteamérica, como Bison priscus gigas, desaparecieron
sin descendencia o dieron lugar al actual
bisonte americano (Bison bison).
Ornimegalonyx oteroi.
Arredondo, 1954
Cuba. Es un género extinto de búho gigante. Conocida como búho
gigante cubano o búho corredor gigante. Se cree que es el búho más
grande que haya existido. Vivía en la Isla de Cuba. Está cercanamente
emparentado con las muchas especies del género actual de búho Strix.
Cuando se describió el primer espécimen en la década de 1950, los huesos
eran tan grandes que se describieron como pertenecientes a un ave de la
familia Phorusrhacidae. En 1961, Pierce Brodkorb revisó los hallazgos y
los atribuyó adecuadamente entre los búhos. Sus restos se encuentran
abundantemente a lo largo de la isla, en depósitos de cuevas desde el
periodo Pleistoceno tardío - Holoceno (de hace unos 10.000 ańos) y se
han encontrado por lo menos tres esqueletos casi completos.
Apteribis sp.
Olson & Wetmore, 1976.
Hawái. Valiéndose de plumas
con una antigüedad de entre 700 y 1.100 ańos, pertenecientes a una
especie de ibis hawaiano, extinguida hace ya tiempo, se ha conseguido
determinar la ubicación de esta ave (Apteribis sp.) en el
árbol genealógico evolutivo de las ibis. Estas plumas son el único
plumaje hallado de las aves prehistóricas extintas que habitaron las
islas de Hawái. Encontradas junto con un esqueleto casi completo, las
plumas han conservado la estructura microscópica necesaria para que los
ornitólogos Carla Dove y Storrs Olson, del Museo Nacional de Historia
Natural, dependiente >aqdel Instituto Smithsoniano, en Estados Unidos,
hayan logrado clasificar con certeza a esta ave como pariente cercana de
la Eudocimus albus y la Eudocimus buber. El
análisis de ADN confirma esta clasificación. Las plumas también han
conservado suficiente pigmentación como para permitir a Dove y Olson
determinar que el ave era de un color entre marrón-negro y marfil-beige.
La Apteribis sp. es una de las dos únicas especies
conocidas de ibis incapaces de volar. Ambas están extintas. El esqueleto
de la Apteribis sp. se diferencia tanto de sus ancestros
del continente que la relación del ave con otras ibis sólo se pudo
determinar a través del estudio de sus plumas y el análisis de ADN. Este
hallazgo es muy inusual porque las plumas no se conservan bien, de modo
que a menudo se descomponen antes de que el ave se haya fosilizado.
Raphus cucullatus.Linnaeus,
1758.
Islas Mauricio. El dodo
o dronte es una especie extinta de ave columbiforme de la familia
Raphidae. Era un ave no voladora endémica de las islas Mauricio,
situadas en el océano Índico. El dodo, así como otras aves del océano
Índico, entre ellas el solitario de Rodríguez y el ibis sagrado de
Reunión (Raphus solitarius), estaba relacionada con las
palomas que habían dejado de volar para volverse terrestres.
Por su temprana extinción, es difícil tener
una descripción precisa del dodo. Las que hay se basan por un lado en
las descripciones y dibujos antiguos, y por otro lado en los esqueletos
y restos encontrados,uno de los cuales se preserva hoy en su posible
postura natural. Como adaptación a la vida en la isla, los dodos
perdieron la capacidad de vuelo, y con ello sufrieron también una fuerte
regresión en la musculatura y de sus ligamentos en el esternón, además
de una transformación en el plumaje, que se volvió filamentoso; la cola
se acortó extraordinariamente y quedó provista de solo unas pocas plumas
arqueadas y fijadas débilmente. Partiendo de esto, se puede decir que el
dodo era un ave de aproximadamente un metro de altura, con un peso que
oscilaba entre 13 y 25 kg y de plumaje grisáceo. Su pico era muy largo
(23 cm) y con una punta en forma de garfio que probablemente le permitía
romper las cortezas de los cocos. Sus patas eran amarillas y robustas,
con unas cuantas plumas rizadas en su parte de atrás. Tenía unas alas
muy pequeńas, que unidas a su gran peso y un esternón insuficiente para
soportarlo le hacían incapaz de volar. El ser humano llegó a su hábitat
en el siglo XVII. Las primeras noticias que se tuvieron en Europa del
ave parecen datar de 1574; en 1581 un conquistador espańol llevó un
ejemplar a Europa. Los descubridores portugueses llamaron "dodo"
(«estúpido» en el habla coloquial portuguesa) al ave por su torpeza y la
facilidad con que podía ser cazada (el dodo evolucionó sin ningún
contacto con seres humanos, por lo que no los veía como una amenaza).
También se le ha llamado dronte, una denominación algo más científica.
El último
ejemplar fue visto por última vez en 1662, aunque existe un avistamiento
por parte de un esclavo escapado en 1674, y se estima que debió de
existir hasta 1690.
Panthera
leo spelaea.
Goldfuss 1810
Estados Unidos, México y Argentina.
Tal vez sea uno de los mayores
félidos que hayan existido, sólo ligeramente superado
por el "tigre" de dientes de sable suramericano (Smilodon
populator), con el que compartía la misma alzada
(alrededor de 1,20
m),
pero el Smilodon poseía una estructura física más
corpulenta, más musculosa, y por tanto fue más pesado. El
león de las cavernas también fue superado por su antepasado
del
Pleistoceno Temprano y Medio el (Panthera
leo fossilis) con aproximadamente 1,25 m de altura
al hombro y sobre todo por el león americano (Panthera
leo atrox) que alcanzó una altura cercana al 1.30 m.
Se hallaba difundido desde las
Islas Británicas en Occidente hasta el
Turkestán en Oriente. Su
taxonomía ha sido muy debatida: pudo ser una especie
diferente (Panthera spelaea), según algunos una clase
de (tigre)
(Panthera tigris spelaea), pero está muy extendida la
opinión reciente de que era una raza geográfica (lo que en
otros términos es llamado en taxonomía una subespecie) del
león moderno (Panthera leo spelaea), que es la
que se utilizará en este artículo. Los estudios de
ADN han probado, finalmente, que el de las cavernas era
un auténtico león, aunque no estaba emparentado con ninguna
subespecie actual en especial.
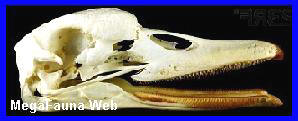
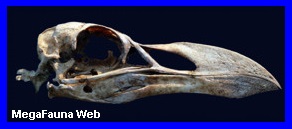
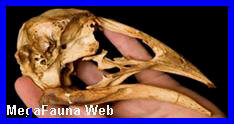
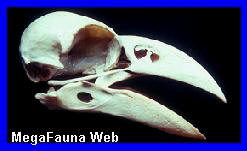
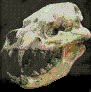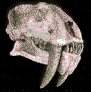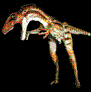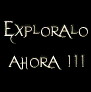
Pygoscelis
adeliae. Hombron
y Jacquinot, 1841.
Cráneo de
pingüino Pygoscelis adeliae, holoceno de Antartida. Imagen;
Dirección Nacional del Antártico
Aspecto del
Pinguino Pygoscelis adeliae. (*).
Antartida.
El orden Sphenisciformes (pingüinos), que comprende una única
familia (Spheniscidae), constituye un grupo de aves con caracteres
altamente derivados. La totalidad de las especies conocidas son aves no
voladoras y con particulares adaptaciones al buceo submarino.
Los pingüinos son las únicas aves vivientes no voladoras
adaptadas al buceo propulsado por las alas. Por ello, sus alas se han
convertido en aletas con huesos fuertemente comprimidos y articulaciones
rígidas que impiden el movimiento independiente de los huesos del ala.
El géneroPygoscelisse alimenta fundamentalmente de
plancton, a diferencias del resto de las especies que comen peces.
Restos fósiles de Pygoscelis adeliaede unos 6.400
ańos de antigüedad, descubierto durante las investigaciones realizadas
en adyacencias de la Base Jubany (caleta Potter, isla 25 de Mayo,
Antártida). Los restos
fósiles descubiertos en estas arenas de playa consisten en huesos de
focas (elefante marinoMirounga sp), huesos y plumas de
pingüinos Adelia (Pygoscelis adeliae) y Papúa (P.
papua) junto a huesos de skúa (Catharacta sp), uno
de los predadores de los pingüinos. Se conocen restos de
Pygoscelis calderensis del Mioceno de Chile.
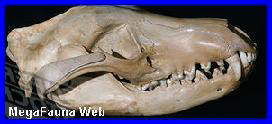
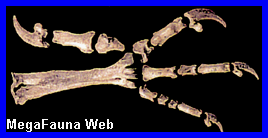
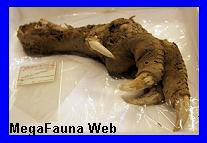
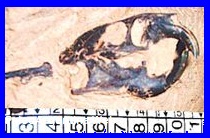
Canis (Dusicyon) avus.
Burmeister, 1864.
Fragmento
mandibular atribuido a Canis Dusicyon
avus. Museo de Miramar.
Esqueleto de
Canis sp en sedimentos Holocenos. Ilustrativo.
Aspecto en vida de
Canis Dusicyon avus.
(*)
Brasil, Chile y Argentina. El presente genero se lo reconoce fosiliferamente solo en el Pleistoceno bonaerense, mientras que en
el Holoceno, se lo encuentra también en la región Patagonica y en el
sector austral Chileno. Tenia la apariencia de un lobo moderno y
pertenece a la fauna que emigro desde el hemisferio norte. Su
alimentación consistía principalmente de pequeńos mamíferos y aves
de zonas abiertas. Si bien su registro es muy escaso y solo se lo
reconoce por restos aislados, es probable que viviera en jaurías
organizadas, y vínculos sociales complejos como las formas
vivientes. En la Provincia de Buenos Aires, se han hallado
asentamientos aborígenes que utilizaban los caninos (colmillos) de
estos, para usos religiosos. La
contextura de Canis Dusicyon era muy parecida a la de un ovejero
alemán. Estudios realizados por los prestigiosos Paleontólogos
Argentinos Walter Berman y Eduardo Tonni, establecieron varias hipótesis
sobre la extinción de este canido, sosteniendo que una de las
posibilidades fue la hibridizacion de esta especie con Canis (Canis)
familiaris, traído de Europa, es decir, que el mismo pudo haber
desaparecido por una cruza con el nuevo representante, pero también
se sospecha de los cambios climáticos y ambientales durante el
siglo XVI.
Los últimos registros de ejemplares de Dusicyon
avus
han sido datados en alrededor de 3000 ańos de antigüedad y coincide
con la gran expansión poblacional de las sociedades
cazadoras-recolectoras de la región.
Dusicyon australis.
Kerr, 1792.
Cráneo y mandíbula deDusicyon australis. Fuente;
sciencephoto. The Falklands Museum.
Ejemplar
taxidermisado
Dusicyon australis. (*)
Islas Malvinas, Argentina.El
lobo austral o zorro antártico, es una especie extinta de mamífero
carnívoro de la familia Canidae endémico de las islas Malvinas,
de dimensiones intermedias entre las del lobo y las de un
zorro grande. Era uno de los dos únicos mamíferos terrestres nativos de
las Malvinas.El guará medía unos 90 cm de longitud corporal desde el
hocico hasta el nacimiento de la cola, que alcanzaba los 30 cm. Según
una de las primeras hipótesis (hoy obsoleta), la presencia del guará en
las Malvinas derivaba de esporádicos desembarcos de canoeros yámana o
manne'ken (aborienes) del archipiélago de Tierra del Fuego. Estas
etnias fueguinas habían logrado domesticar a los culpeos creando el
antiguo perro yagán o perro fueguino. En 2009 un estudio
de ADN confirmó que el pariente vivo más cercano es en realidad el
aguará guazú. El estudio confirmó que ambas especies se separaron hace
alrededor de 6,7 millones de ańos. En 2013, un estudio de
ADN mitocondrial de diversas especies de cánidos sudamericanos (vivos y
extintos) determinó que el guará habría divergido hace 16 000 ańos deDusicyon avus, una especie similar que habitó en la Patagonia
hasta hace unos 3000 ańos. El registro histórico más antiguo sobre este
animal se dio en el viaje del capitán John Strong en 1690 y era aún
bastante común en el tiempo en que Charles Darwin visitó las islas
Malvinas en 1833. Igualmente Darwin predijo la extinción del zorro.
También se lo conoce como Canis antarcticus.
Eutatus seguini.Gervais, 1867.
Esqueleto completo
de Eutatus seguini, en el Museo Paleontológico de Salto.
Coraza
parcialmente completa de Eutatus. Arroyo Carnero,
Prov. de Córdoba. Museo de la Univ. de Córdoba.
Aspecto del
actual Tatu Carreta, que comparte semejanzas con Eutatus seguini.
Por
Carlos C. Wiedner.
Argentina. Fue
un dasipodido muy común en el territorio bonaerense, que recuerda en
cierta forma con el Tatu Carreta. Eutatus
seguinifue descrito originalmente en el ańo 1867 por el
paleontólogo francés François Louis Paul Gervais. El material tipo
son restos adquiridos en el ańo 1871 por el Muséum National d'
Histoire Naturelle de París, los que fueron colectados por
François Seguin. Su registro fosilífero abarca
desde el Pleistoceno inferior (2 millones de ańos) hasta el
Holoceno temprano ( 8 mil ańos), encontrándose en algunos casos,
asociados a restos óseos de otros animales por actividad antropica,
es decir, que han presentado manipulación por antiguos grupos
humanos. Al igual que los armadillos, pose un escudete de placas óseas
en su cráneo, conocido también como escudete cefálico. Su coraza
dorsal es robusta, y poseía unas 33 bandas móviles que cubrían
las dos terceras partes del caparazón. En cada mandíbula poseía
de 9 a 10 dientes, los cuales utilizaba para comer carne en
descomposición, huevos, larvas, caracoles y algunos tallos. Construían
grandes galerías subterráneas, con cámaras amplias para proteger
sus crías. Su cráneo era alargado. Sus patas eran cortas y
robustas, protegidas por unas enormes falanges ungueales o garras,
las cuales, eran justamente utilizadas para cavar.
Pontoporia
(Stenodelphis)
blainvillei.
Norman & Fraser, 1938.
Mariano Magnussen
Saffer con un esqueleto de Delfín del Plata o Franciscana
(Pontoporia
blainvillei) en el Museo
Municipal de Ciencias Naturales Punta Hermengo de Miramar.
Stenodelphis.
Ilustración de Carlos C. Wiedner.
Argentina.
Los Odontoceti son los cetáceos que poseen dientes. En general, los
dientes son todos muy similares entre sí, cónicos, puntiagudos, con una
sola raíz, y con una función parecida. Algunas especies tienen más de
100 dientes en cada hemimandíbula, mientras que otras, poseen sólo dos.
Los odontocetos son conocidos desde el Oligoceno, mientras que en la
Argentina datan recién del Mioceno temprano (hace alrededor de 20
millones de ańos). La familia Pontoporiidae incluye a formas que están
adaptadas a vivir en ambientes de agua dulce. En la Argentina, dicha
familia data del Mioceno tardío (hace algo más de 5 millones de ańos).
En la actualidad, la especie más característica es Pontoporia
blainvillei (o delfín del plata), presente desde el Pleistoceno
tardío-Holoceno en la Argentina. Pontoporia es un delfín pequeńo, de
casi 1,5 metros de largo, con el hocico delgado y largo. Habita en aguas
saladas cerca de la costa, incursionando en ríos aguas arriba. Los
odontocetos vivientes que habitan o visitan los mares de la Argentina se
agrupan en cinco familias: Phocoenidae, Physeteridae, Pontoporiidae,
Delphinidae y Ziphiidae (las tres últimas también con representantes en
el Pleistoceno- Holoceno).
Hippopotamus
antiquus.Desmarest,1822.
Espańa, Inglaterra y Grecia. Es una
especie
extinta de hipopotámido que vivió en amplias zonas de
Europa durante el
Pleistoceno y que se extinguió justo antes de la última
glaciación. El hábitat deH. antiquus comprendía
desde la
Península Ibérica a las
Islas Británicas y desde la cuenca del
Rin
hasta
Grecia.En cuanto a sus dimensiones, el hipopótamo
europeo tenía un tamańo similar a otra especie extinta,
Hippopotamus gorgops y más grande que el actual
hipopótamo común (Hippopotamus
amphibius). Se cree que el hipopótamo europeo
apareció hace alrededor de 1,8 millones de ańos, durante el
Pleistoceno Medio. H. amphibiusvivió igualmente
en el viejo
continente, pero no es considerado "europeo", por sus
orígenes
africanos. El también extinto
Hippopotamus creutzburgi(hipopótamo enano
cretense), es considerado un pariente del europeo que
evolucionó a través del proceso de
enanismo insular en la isla de
Creta.
Cygnus
falconeri.
Parker,
1865.
Malta y Sicilia. Era un muy grande Siculo-Maltés
cisne sabido de
Pleistoceno medio - Holoceno. Sus dimensiones se describen como exceder los de
la vida
Cisne mudo por una mitad,
cuál daría una longitud de la cuenta-a-cola de cerca de 190-210 centímetro
(basado en 145-160 centímetro para C. olor).
Habría sido más alto, aunque no más pesado, que la región
elefantes enanos. Debido a su tamańo, pudo haber sido flightless.
Llegó a estar extinto antes del aumento en
humano actividad en la región, así que su desaparición se
piensa para haber resultado de extremo
fluctuaciones del clima o la llegada del superior
depredadores y
competidores. El expediente del fósil del género Cygnus es absolutamente
impresionante, aunque la asignación a los subgéneros es a menudo
tentativa; según lo indicado arriba, por lo menos las formas tempranas
pertenecen probablemente a C. olor- Linaje meridional del
hemisferio, mientras que los taxa Pleistocenos de Norteamérica serían
puestos adentro Olor. Un número de especies prehistóricas se han
descrito, sobre todo del hemisferio norteńo. Entre ellos estaba el
gigante
Siculo-Maltés
C. falconeri cuál era más alto (sin embargo no más pesado) que
los elefantes enanos locales contemporáneos (Falconeri
de Elephas).
Los cisnes fósiles supuestos Bilinicus de “Cygnus” y
Herrenthalsi de “Cygnus” era, respectivamente, a
cigüeńa y algún pájaro grande de la afinidad desconocida
(debido al mal estado de la preservación del material
referido). Atavus de Anserse coloca a veces adentro
Cygnus.
Pinguinus
impennis.Linnaeus. 1758.
Norteamérica
y Europa. Es una
especie
extinta de
ave Charadriiforme de la
familia Alcidae.
Fue la más grande de las
alcas. A diferencia de las especies actuales de alcas, la gigante
carecía de la capacidad de vuelo, aunque era una buena nadadora y
buceadora. Los ejemplares adultos tenían alrededor de un
metro de altura, unos 78 centímetros y llegaban a pesar 5 kilogramos. El
plumaje era negro en las alas y la espalda, cuello y cabeza. A los lados
de ésta destacaban dos manchas blancas, del mismo color que el abdomen. Su
incapacidad para volar y lo apetitoso de sus huevos (13 centímetros de
longitud y 400 gramos de peso) y carne las hicieron un apreciado
alimento y unas presas perfectas ya en la
prehistoria, como demuestran varios yacimientos
paleolíticos. Fue sometida a una caza despiadada: de otra parte,
eran aves confiadas en exceso, que se dejaban capturar con gran
facilidad. A finales del
siglo
XVI el alca gigante ya había desaparecido de la Europa continental y
en
América del Norte sólo abundaba al norte de
Nueva York. Los naturalistas del
siglo XVIII describen su sabor como atroz, pero parece que los
marineros no tenían un paladar tan exquisito y paraban a menudo durante
sus viajes para aprovisionarse de su carne y, sobre todo, de sus huevos.
Cuando
Linneo nombró la especie, inicialmente como Alca impennis, en
1758, el alca gigante era un animal sumamente raro en Europa, incluso en islas
del
mar del Norte donde un siglo antes abundaba. En
1790 se
capturó un ejemplar en
Kiel, lo
que causó gran extrańeza por ser el único visto en el
mar Báltico en ańos. Hacia
1800, la
especie ya se había extinguido en Norteamérica y su distribución se
reducía a Islandia.
Palaeopropithecus
ingens.G. Grandider,
1899.
Madagascar. Es un
género
extinto de
primates
estrepsirrinos, uno de los tres recientemente
extinguidos de
lémures perezosos encontrados en
Madagascar, y que estaba estrechamente relacionado con
algunas especies de
lémur que se encuentran en la isla en la actualidad. Se
conocen tres especies dentro de este género, P. ingens,
P. maximus y
P. kelyus. Recientes
dataciones por radiocarbono indican que podrían estar
todavía vivos alrededor del 1500 EC.
Se cree que Palaeopropithecus ingenspodría ser el
primate de gran tamańo conocido en algunas leyendas
malgaches como «tretretretre», y del que hablaba
Étienne de Flacourt, gobernador francés de Madagascar a
mediados del siglo XVII, en su L’Histoire de le Grande
Île de Madagascar (1658), que algunos autores
identificaron inicialmente con otro género de lémures
extintos, MegaladapisDurante mucho tiempo se creyó que que
Palaeopropithecuseran criaturas acuáticas, que nadaban
con sus ojos, oídos y fosas nasales justo por encima del
agua. Sin embargo esta teoría estaba basada en un resto
postcraneal mal atribuido. Charles Lamberton refutó este
hecho en 1957. Entonces, los fósiles antes asignados a
cráneos de otras especies se reasignaron a
Palaeopropithecus. Pruebas postcraneales indican un
estilo de vida muy arbóreo con una locomoción de suspensión.
Palaeopropithecus
junto con Babakotia radofilai.
,
Archaeoindris y
Mesopropithecus, forma la familia
Palaeopropithecidae, conocidos con el
nombre de lémures perezosos.
Thylacinus cynocephalus.Harris, 1808.
Australia y Nueva
Guinea. También conocido como lobo de Tasmania,
tigre de Tasmania y tilacín, fue un
marsupial
carnívoro originado en el
Holoceno. Era nativo de
Australia y
Nueva Guinea y se cree que se
extinguió en el
siglo XX. Se trataba del último miembro viviente de su
género (Thylacinus),
viviendo los otros miembros en tiempos prehistóricos a
partir de principios del
Mioceno.
El lobo marsupial se extinguió en el
continente australiano miles de ańos antes de la llegada
de los
colonos europeos, pero sobrevivió en
Tasmania junto con otras
especies endémicas, incluyendo el
diablo de Tasmania. Generalmente suele culparse de su
extinción a la caza intensiva, incentivada por recompensas,
pero podrían haber contribuido otros factores, como por
ejemplo las
enfermades, la introducción de los
perros, o la ocupación de su
hábitat por los humanos-
Aún cuando se lo considera oficialmente extinto, todavía hay
quienes dicen haberlo visto. Como los
tigres y
lobos del
Hemisferio Norte, de los cuales heredó dos de sus
nombres comunes, el lobo marsupial era un
depredador alfa. Siendo un
marsupial, no tenía relación con estos mamíferos
placentarios, pero debido a la
evolución convergente, presentaba la misma forma general
y las mismas adaptaciones. Su pariente vivo más próximo es
el
diablo de Tasmania.
Harpagornis moorei.Haast, 1872.
Nueva Zelanda. Es una
especie
extinta de
ave
falconiforme de la
familia Accipitridae que habitaba en
Nueva Zelanda. Después de la extinción de los
teratornítidos (grandes aves carrońeras de la Era del
Hielo, que vivieron en
América y eran parientes de los
cóndores), el harpagornis se convirtió en la mayor
ave rapaz del mundo. Se cree que los
maoríes lo llamaban pouakai o hokioi. Era
una rapaz enorme; las hembras de harpagornis pesaban entre
10 y 15 kg y los machos, de 9 a 10 kg. Tenían una
envergadura de alas de alrededor de 3 metros, la cual es
poca si se considera el tamańo que tenían, pero esto les
permitió cazar en áreas boscosas. Ésta ave
predadora se alimentaba de otras especies de
aves no voladoras, principalmente de las
moas. Los primeros habitantes que se establecieron en
Nueva Zelanda, no sólo cazaron las moas sino también las
harpagornis, llevando a ambas especies a su extinción. Esta
ave fue clasificada por
Julius von Haast, quien la llamó Harpagornis moorei,
después de que
George Henry Moore encontrara huesos de esta especie en
sus tierras.
Aptornisdefossor.Owen, 1871.
Nueva Zelanda. Es un
género
extinto de
aves
gruiformes, el único de la
familia
Aptornithidae, que incluye dos especies estrechamente
relacionadas
endémicas de
Nueva Zelanda,
Aptornis otidiformis (Isla
del Norte) y
Aptornis defossor (Isla
del Sur). En vida, eran aves masivas, menores que una
moa (con las que en un principio fueron confundidos),
con un gran pico curvado hacia abajo y patas fuertes. Eran
incapaces de volar ya que tenían las alas muy reducidas,
menores incluso que las de el
dodo y con un
carpometacarpo reducido de un modo único. Las dos
especies conocidas se diferenciaban sobre todo en el tamańo,
siendo la especie de la Isla del Norte menor que la del Sur;
su color en vida se desconoce. Los fósiles se han encontrado
en las zonas más áridas de Nueva Zelanda y solo en las
tierras bajas. Richard Owen, que describió las dos especies,
especuló en que podrían ser
omnívoras y el análisis
isotópico de los huesos lo confirma. Los niveles de
13C y 15N de dos especímenes de Aptornis
otidiformis comparados con los mismos valores en la moa,
en
Chenonetta finschi(Anatidae)
y en
Aegotheles (ave insectívora) sugieren que
Aptornis comía especies situadas más arriba en la
cadena trófica que los insectívoros.
Se cree que comían grande
invertebrados,
lagartos,
tuátaras e incluso
pájaros. Fueron cazados por los colonizadores
polinesios y sus huevos depredados por
perros y
ratas. Se extinguieron antes de la llegada de los
exploradores
europeos.
Chelychelynechen quassus.Olson & James, 1991.
Hawái. El Chelychelynechen, que
quiere decir pato de mandíbula de tortuga, tenía un pico grande y pesado
como el de las tortugas, mientras que los otros dos géneros, el
Thambetochen y el Ptaiochentenían los lados del pico como
sierras conocidas como pseudodientes. Ninguna especie podía volar,
siendo su tamańo bastante grande, con un peso de entre 4 y 7,5 kg.
fueron un grupo de aves que se
extinguieron de forma extrańa, un tipo de
pato que
vivió en las islas más grandes de
Hawái,
menos en la isla mayor, en el Pacífico. Fueron los mayores
herbívoros de estas islas en los últimos 3 millones de ańos, hasta
que se extinguieron después del asentamiento humano. La extrańa forma y
talla de los moa-nalo se atribuye a su papel dentro de la ecología de la
prehistórica Hawái`i. Un estudio de los coprolitos (excrementos fósiles)
del Thambetochen Chauliodous encontrados en la cueva de Pu`u Naio en las
tierras bajas de Maui mostró que eran comedores de hojas, especialmente
de hoja de helechos (posiblemente
Asplenium nidus o
Dryopteris wallichiana). Esta conclusión explica la forma
característica de sus picos
Oceanía.
Son una
subfamilia de
cocodrilos de
Australia y de
Oceanía
que se ha extinguido recientemente. Aparecen por primera vez en el
registro fósil en el
Eoceno
en Australia, y sobrevivieron hasta el
Pleistoceno en Australia, tras la llegada de los cocodrilos del
género
Crocodylus, que hoy en día aún pueblan el continente; los del
género
Mekosuchus perduraron hasta la llegada del ser humano a las
islas del Pacífico, como
Fiji,
Nueva Caledonia y
Vanuatu.
Fueron un grupo muy diverso. La mayoría de los cocodrilos de la
Australia del Cenozoico se cree que fueron una radiación endémica de
mekosuquinos. A pesar de su diversidad ecológica y geográfica, muchos de
estas especies de cocodrilos mekosuquinos se extinguieron en el
continente australiano, no mucho después de la llegada de Crocodylus,
en algún momento hacia el final del Plioceno. A finales del Pleistoceno,
tras la llegada de los humanos y los
dingos, todos ellos habían desaparecido. En muchas de las islas del
Pacífico Sur, sin embargo, los mekosuquinos persistieron. Algunos, como
el
Mekosuchus de
Nueva Caledonia, Mekosuchus inexpectatus, sobrevivieron en
tiempos históricos.
Daubentonia
robusta.Lamberton, 1934.
Nueva Zelanda. Es una
especie de
primate
estrepsirrino que se
extinguió hacia 1920; era próximo al
aye-aye.En 2007, los restos conocidos de esta
especie eran 4
incisivos, una
tibia y material postcraneal. Fósiles de esta especie han sido
encontrados al este y sureste de
Madagascar fuera de las zonas en las que habita el aye-aye, animal al que se cree que era muy similar
morfológicamente, sólo que 2-2,5 veces más grande basándose
en las medidas de una mandíbula y del incisivo.
Dinornis giganteus.Owen,
1844.
Nueva Zelanda.
El moa gigante de la isla Sur
es una especie extinta de aves dinornitiformes de la
familia Dinornithidae. Eran aves no voladoras con un
esternón sin quilla. También tienen un paladar diferente. El
origen de estas aves se vuelve más claro ya que se considera
ahora que evolucionaron de aves voladoras que perdieron la
capacidad de vuelo tras su llegada a las islas de Nueva
Zelanda. Vivía en la isla Sur y su
hábitat eran los matorrales, dunas, pastizales y bosques en
tierras bajas. En
1878. Sir Richard Owen,
una de las mayores autoridades de Europa en osteología,
identifica, a partir de un único fragmento de hueso de una
de sus patas al Moa Gigante de Nueva
Zelanda, un
ave que desapareció en el
periodo Holoceno,
es decir, en el que vivimos actualmente, sólo que el hombre
fue uno de los más directos responsables de su extinción.
Cuando los maoríes
llegaron a Nueva Zelanda hace 1.000 ańos comenzó la caza de
este enorme pájaro que llegaba a medir casi 3 metros de alto
y que desapareció definitivamente, se cree, en el mismísimo
siglo XIX.
Aguirre, M. L. 1990.
Asociaciones de moluscos bentónicos marinos del cuaternario
tardío en el noroeste bonaerense. Ameghiniana 27: 161-177.
Alberdi, M.T.; Prado, J.L.
1995b. Los Équidos de América del Sur. In Evolución
climática y biológica de la región pampeana durante los
últimos cinco millones de ańos. Un ensayo de correlación con
el Mediterráneo occidental (Alberdi, M.T.; Leone, G.; Tonni,
E.P.; editores). Museo Nacional de Ciencias Naturales,
Consejo Superior de Investigaciones Científicas,
Monografías, p. 295-308. Madrid.
Ameghino, F. 1887.
Enumeración sistemática de las especies de mamíferos fósiles
coleccionados por Carlos Ameghino en los terrenos eocenos de
Patagonia austral y depositados en el museo de La Plata.
Boletín del Museo de La Plata 1: 1-26.
Benton,
Michael. (1992). Dinosaurios y otros animales prehistóricos.
Ediciones Lrousse Argentina S.AI.C.
Berman, W. D. 1994. Los
carnívoros continentales (Mammalia, Carnivora) del Cenozoico
en la provincia de Buenos Aires. Ph.D. dissertation,
Universidad Nacional de La Plata, La Plata, Argentina, 413
pp.
Berman, W.
D. y Tonni, E. P. 1995. Algunas consideraciones sobre Canis
(Dusicyon) avus (Carnívora: Canidae) en el Pleistoceno
tardío y Holoceno de la provincia de Buenos Aires (Republica
Argentina).
I Jorn. Geol. Prov. Bs. As., Tandil, 11-15
nov. de 1985, res.: 215-216.
Berman, W. D. y Tonni, E.P.
1988. Canis (Dusicyon) avus (Carnívora, Canidae) en el
Pleistoceno tardío y Holoceno de la provincia de Buenos
Aires. Aspectos sistemáticos y bioestratigraficos
relacionados. Ameghiniana, 24 (3-4): 245- 254. Buenos Aires.
Cabrera, A., 1957. Catalogo
de los mamíferos de América del Sur. I. Rev. Mus. Cs. Nat.
“B. Rivadavia”, Zool. 4(1); 1-307.Bs.As.
Francesco C.G. y Zarate, M.A.1999.
Análisis tafonomico de Littoridina Souleyet, 1852 (Gastropoda:
Hydrobiidae) en perfiles holocenos del rió Quequen grande
(Prov. Buenos Aires): significativo paleobiologico y
paleoambental. Ameghiniana 36 (3): 297-310 Bs.As.
Francisco
J. Prevosti Ubilla, M., & Perea, D. (marzo de 2009). «Large
extinct canids from the Pleistocene of Uruguay: systematic,
biogeographic and paleoecological remarks». Historical
Biology: A Journal of Paleobiology(en inglés).
Vol. 21, N° 1-2.
Consultado el 2 de julio de 2012. «79-89».
Fidalgo, F. y Tonni, E.P.
1983. Geología y paleontología de los sedimentos encausados
del pleistoceno tardío y holoceno de Punta Hermengo y arroyo
Las Brusquitas (Partido de General Alvarado y General
Pueyrredon, Provincia de Buenos Aires). Ameghiniana 20
(3-4): 281-296.
Firinati, E.A. y Aliotta,
S.A. 1995. Análisis tafonomico de conchillas en cordones
holocenos, Bahía Blanca, Argentina. 4 Jorn. Geológicas y
geofísicas bonaerenses (Junin) Actas 1: 89 – 97.
Frenguelli, J. 1920. Los
terrenos de la costa atlántica en los alrededores de Miramar
(prov. Bs.As.) y sus correlaciones. Bol. Acad. Nac. Cienc.
Cordoba 24: 325-385.
Francesco C.G. y Zarate,
M.A.1999. Análisis tafonomico de Littoridina Souleyet, 1852 (Gastropoda:
Hydrobiidae) en perfiles holocenos del rio Quequen grande (Prov. Buenos
Aires): significativo paleobiologico y paleoambental. Ameghiniana 36
(3): 297-310 Bs.As.
Gaillard, C. y Castellanos,
Z.A. 1976. Fauna de agua dulce de la republica Argentina:
volumen 15: Gasterópodos; fascículo 2 :Hydrobiidae. Fecil
1:39 p.
Palanca, F. Y Politis, G.
1979. Los cazadores de fauna extinguida de la provincia de
Buenos Aires. Prehistoria Bonaerense, pp. 71-91, Olavaria.
L. Miotti, M. Salemme y J.
Rabassa (2000): «Secuencia radiocarbónica de Piedra Museo»,
en: Guía de campo de la visita a las localidades
arqueológicas. Taller internacional "La colonización del
sur de América durante la transición Pleistoceno/Holoceno",
págs. 83-87.
Lavallee, D. 1990. La
domestication animale an amerique du Sud. Le point des
connaissances. Boletín del Instituto Francés de Estudios
Andinos. 19 (1): 169-176. Buenos Aires.
Magnussen Saffer,
Mariano. 2015. Los
mamíferos, aves, reptiles y anfibios que coexistieron con el
hombre en el holoceno pampeano.
Paleo Revista Argentina de
Paleontología. Boletín Paleontológico. Ańo XIII. 128: 28-29.
Menegaz, A.N.;
Ortíz Jaureguizar, E. 1995. Los Artiodáctilos. In Evolución
biológica y climática de la región Pampeana durante los
últimos cinco millones de ańos. Un ensayo de correlación con
el Mediterráneo occidental (Alberdi, M.T.; Leone, G.; Tonni,
E.P.; editores). Museo Nacional de Ciencias Naturales.
Consejo Superior de Investigaciones Científicas,
Monografías, p. 311-337.
Madrid.
NOVAS, F. 2006. Buenos Aires hace un millón
de ańos. Editorial Siglo XXI, Ciencia que Ladra. Serie
Mayor.
Olrog C. C. y Lucero M. M.
1981. Guía de los mamíferos argentinos. Fundación Miguel
Lillo. Tucumán. Argentina.
Palanca, F; Daino, L; y
Berbassat, E. 1979. Los cazadores de la fauna extinguida de
la Provincia de Buenos Aires. Prehistoria bonaerense, pp.
71-91. Olavaria.
Politis, G. 1984.
Investigaciones arqueológicas en el área Interserrana
Bonaerense. Etnia 32: 7-52. Olavaria.
Reig, O. A. 1964. Roedores
y marsupiales del partido de General Pueyrredon y regiones
adyacentes (provincia Buenos Aires, Argentina) Publ. Mus.
Munich. Cienc. Nat. Y Trad. Mar del Plata. 1 (6): 297-352.
Rodrigues, P.H.; Prevosti, J.F.;
Ribeiro, A.M.; Ferigolo, J. (2004) Novos materiais de
carnívora para o Pleistoceno do Estado do Rio Grande do Sul,
Brasil. Revista Brasileira de Paleontologia, v.7, n.1,
p.77-86, 2004.
Tambussi P.
C. y Tonni E.P. La distribución del género Rhea (Aves:
Rheiformes) en el Pleistoceno tardío-Holoceno de la Región
Patagónica.
I Jorn. Arg. Paleont. Vert. res., 11; 1984
Ver mas
bibliografía utilizada para hacer la presente Pagina;
AQUÍ.
Ver bibliografía de divulgación sugerida; AQUÍ.
Nuestro sitio Web posee una amplia
y completa información sobre geología, paleontología,
biología y ciencias afines. Antes de realizar una
consulta por e-mail sobre algunos de estos temas,
utilice nuestro buscador interno. Para ello utilice
palabras "claves", y se desplegara una lista de
"coincidencias". En caso de no llegar a satisfacer sus
inquietudes, comuníquese a
grupopaleo@gmail.com