Los mamíferos dientes de sable y su
paso en la historia biológica.
Por
Mariano Magnussen Saffer. Grupo Paleo. Museo
Municipal Punta Hermengo de Miramar. Fundación
Argentavis.
marianomagnussen@yahoo.com.ar. Tomado de; Magnussen Saffer,
Mariano. 2015. Los
mamíferos dientes de sable y su paso en la historia
biológica. Paleo Revista Argentina de
Paleontología. Boletín Paleontológico. Ańo XIII. 132: 13-18.
Desde hace unos 35 millones de ańos hasta hace 12
mil ańos antes del presente (Pleistoceno superior),
vivieron en distintas edades geológicas y distintos
continentes un grupo muy peculiar de mamíferos, los
cuales desarrollaron como parte de un proceso
evolutivo increíble, la presencia de dos caninos
(colmillos) superiores altamente desarrollados.
 |
Estas especies adquirieron sus
característicos dientes de sable de forma
totalmente independiente, aun teniendo poco
parentesco, geografía y edad geológica. La
morfología de los dientes es un excelente
ejemplo de evolución convergente o evolucion
paraslela en otros casos, ya que ha ocurrido
repetida e independientemente en por lo
menos cuatro grupos diferentes de mamíferos:
los macairodontes (Machairodontinae), una
subfamilia de los felinos que incluye al
famoso Smilodo s.p.; los nimrávidos (Nimravidae),
una familia emparentada con los felinos,
pero diferente; y finalmente, un par de
casos aislados dentro de los creodontos (que
no son considerados carnívoros auténticos) y
los boriénidos, una familia extinta de
marsupiales sudamericanos. |
El
consenso general de acuerdo a su dentadura es que se
usaban para cazar denotando con ello una
alimentación carnívora, la forma exacta en la que se
usaban ha sido motivo de debate desde la década de
1880, cuando el Smilodon fue descrito por
primera vez.
Algunos paleontólogos creen que el propósito
principal era el de agarrar y sujetar presas
grandes. La evidencia sin embargo no da suficiente
respaldo a esta teoría, ya que se ha demostrado que
unos dientes tan largos usados de esa forma podrían
romperse con facilidad, y los cráneos fósiles con
los dientes rotos son raros.
Una
hipótesis más aceptada sugiere que los dientes de
sable se usaban para dar mordiscos en la garganta o
el abdomen de presas grandes, o para causar heridas
punzantes profundas que desangrarían al animal.
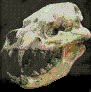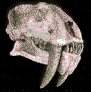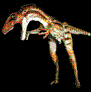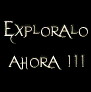
Hoplophoneus.
Cope,
1874.
Vivieron en Norte América durante el
Eoceno-Oligoceno (hace 38 - 33,3 millones de ańos).
Existiendo durante aproximadamente
4,7 millones
de ańos.
 |
Hoplophoneus
era similar a los félidos, aunque no lo
eran. La estructura de los huesos del oído
interno son diferentes; los félidos tienen
una estructura externa denominada bulla
auditiva separada por un tabique en dos
cámaras; los nimrávidos carecen de bulla.
Poseían además un surco en la mandíbula
inferior donde se acomodaban los largos
caninos en forma de sable de la mandíbula
superior. Tenía el tamańo de un leopardo,
con un cuerpo robusto y patas cortas.
|
El
espécimen más grande que se conoce fue examinado por
Sorkin (2008) para establecer su masa corporal, la
cual se estimó en 160 kilogramos.
Paramachairodus.
Pilgrim, 1913.
Paramachairodus
(alternativamente
Paramachaerodus)
es un
género extinto de gato
dientes de sable
de la subfamilia
Machairodontinae, que era
endémica en
Europa y Asia
durante el
Mioceno tardío
15-9 Ma.
|
 |
Paramachairodus es uno de los gatos
dientes de sable
verdaderos
conocidos
más antiguos.
Se
descubrió
una
gran cantidad de fósiles
en el
Cerro de los Batallones,
un
yacimiento de fósiles
del Mioceno
tardío, cerca de
Madrid, Espańa.
Dos
especies
de leopardo
son conocidos,
Paramachairodus
ogygia
y
Paramachairodus
orientalis.
Una
tercera
especie,
Paramachairodus
maximiliani,
se considera
sinónimo de
Paramachairodus
orientalis
por la mayoría de
autores |
Los
animales tenian 58
centímetros
de alto en
el hombro, similar a
un leopardo,
pero con un cuerpo
más flexible.
La forma de sus
extremidades sugiere que puede
haber sido un
escalador
ágil, y podría haber
cazado
presas relativamente grandes.
Metailurus.
Zdansky, 1924.
Es
un género extinto de felino dientes de sable,
perteneciente a la tribu Metailurini, que vivió en
América del Norte, África, Europa y Asia desde el
Mioceno al Pleistoceno, viviendo desde hace 9
millones de ańos hasta hace 11,000 ańos.
Metailurus
fue nombrado por Zdansky (1924). Fue asignado a la
familia Felidae por Carroll (1988).
Metailurus
es conocido como el felino "diente de sable falso"
debido que no poseía verdaderos dientes de sables,
sino que tenía una especie de mezcla entre los
caninos alargados, delgados y en forma de cuchillo
de los verdaderos dientes de sable (Machairodontinae)
y los caninos más cortos y cónicos de los felinos
actuales.
Los
caninos son mayores incluso que los del leopardo
nebuloso pero significativamente más cortos que los
de los dientes de sable verdaderos y más cónicos que
aplanados. Debido a que muchos de los fósiles de
Metailurus son fragmentarios, el número de
especies es objeto de debate
Machairodus.
Kaup, 1833
Es
un género extinto de mamíferos felinos de la
subfamilia Machairodontinae. Eran félidos dientes de
sable que vivieron en Europa, Asia, África y
Norteamérica, aproximadamente desde hace 12 millones
de ańos a 200 mil ańos
Las
especies fósiles asignadas al género Machairodus
fueron divididas por Turner en dos grupos de
desarrollo evolutivo - M. aphanistus y el "Nimravides"
catacopis de Norteamérica representan el
grado más primitivo, y M. coloradensis y
M. giganteus (Amphimachairodus giganteus)
representan el grado más derivado.
Las
características del grado más avanzado incluyen el
relativo alargamiento del antebrazo y el
acortamiento de la región lumbar de la columna que
se parecía a la de los actuales felinos panterinos.
Estas tendencias serían más desarrolladas en
Homotherium, el cual se cree que evolucionó de
Machairodus.
Sus
dientes estaban bien arraigados a su boca y no eran
demasiado delicados, a diferencia de la mayoría de
los felinos dientes de sable de la época, que tenían
colmillos extremadamente largos que sobresalían de
la boca. Los colmillos de Machairodus, sin
embargo, eran capaces de encajar cómodamente en su
boca aún siendo largos y efectivos para la caza.
Dinofelis.
Zdansky 1924.
Es
un género de felinos dientes de sable que pertenece
a la tribu Metailurini. Estaban muy extendidos en
Europa, Asia, África y América del Norte, desde hace
5, hasta alrededor de 1,2 millones de ańos
(principios del Plioceno y comienzos del
Pleistoceno).
|
 |
Los fósiles del Dinofelis y los
huesos se han encontrado en Sudáfrica, junto
con babuinos que posiblemente cazaba.
Los huesos de varios ejemplares de
Dinofelis y babuinos fueron encontrados
en una trampa natural. Dinofelis
puede haber entrado en el lugar para
alimentarse de los animales capturados o
simplemente penetró al sitio y no fue capaz
de escapar de nuevo. |
Varios sitios de fósiles de Sudáfrica parecen
demostrar que Dinofelis pudo haber cazado
Australopithecus afarensis y Paranthropus,
ya que albergaban restos fosilizados de Dinofelis,
estos homínidos y otros animales grandes de la
época.
Tenían un tamańo intermedio entre un leopardo y un
león modernos, la mayoría poseía el tamańo de un
jaguar (70 cm de altura y hasta 120 kg de peso), de
tamańo, de gran alcance, que tenía dos dientes de
sable prominentes. Dos especímenes fueron examinados
por Legendre y Roth para calcular la masa corporal.
En el primer ejemplar se estimó un peso de 31,4 kg.
El segundo se estimó con un peso de 87,8 kg
Homotherium.
Fabrini, 1890.
Es
un género de felinos fósiles que vivieron entre hace
5 millones y 10.000 ańos, durante los periodos
Plioceno y Pleistoceno, conocidos popularmente como
dientes de sable.
En
el momento de su mayor expansión habitaron en la
mayor parte de África, Eurasia, Suramérica y
Norteamérica. Tan vasta distribución y el hallazgo
de cientos de restos en numerosas localidades han
permitido la identificación de muchas especies,
siendo las más conocidas el Homotherium serum
de América del Norte y la especie europea
Homotherium latidens. La especie H. serum
fue una de las últimas de dientes de sable en
desaparecer, junto con las de otro género más
popular, Smilodon.
 |
Los largos colmillos típicos de los
macairodontes están presentes, aunque ni de
lejos tan desarrollados como en Smilodon
o Megantereon, sino que son mucho más
cortos, curvos (por lo que a veces se llama
a los homoterios "felinos - cimitarra") y
finos.
Cuando el animal mantenía la boca cerrada,
sólo se les veía la punta. Siguiendo con la
tónica general del grupo, los homoterios
también tenían los cuartos delanteros muy
desarrollados, con el fin de vencer la
resistencia de las presas de que se
alimentaban antes de clavarles los colmillos
en el cuello. |
Los
hallazgos de varios esqueletos juntos, en ocasiones
acumulados durante generaciones en un mismo lugar, y
entre los que se encuentran sujetos con fracturas
curadas y enfermedades, sugieren que al menos las
especies más tardías vivían en manada y cooperaban
para cazar.
Megantereon.
Cuvier, 1824.
Es
un género extinto de felino dientes de sable
perteneciente a la subfamilia Machairodontinae.
Este mamífero tenía un tamańo similar al leopardo
actual, pero era más robusto y musculoso, y poseía
unos colmillos superiores alargados y ligeramente
curvos que pudo emplear en la caza de presas tipo
mastodonte. Vivió en Eurasia, África y Norteamérica
desde el Mioceno Superior hasta el Pleistoceno
Medio.
 |
A día de hoy no existe consenso entre los
científicos sobre el lugar de origen de este
género.
Se tienen evidencias fidedignas de su
presencia en África desde hace 3,6 millones
de ańos, durante el Plioceno medio, pero
existen restos asiáticos de dudosa
clasificación que podrían adelantar su
existencia hasta el Mioceno. |
En
norteamérica los restos más antiguos conocidos datan
de unos 4,5 millones de ańos, pero algunos autores
no consideran este registro perteneciente al género
Megantereon. En Europa el registro más
antiguo del género data, según autores, entre 3,5 y
2,5 millones de ańos. Este género es considerado por
algunos el ancestro de Smilodon.
Xenosmilus.
Martin, et al, 2000.
Es
un género extinto de mamífero macairodontino, es
decir un felino diente de sable. Dos especimenes
casi intactos fueron hallados por recolectores
aficionados de fósiles en 1983 (1981 según algunas
fuentes) en las minas de calizas de Haile en el
Condado de Alachua, Florida.
En
1994 los fósiles fueron examinados, y se decidió que
pertenecían a un nuevo género de félido. Los fósiles
datan entre 1.8 a 0.3 millones de ańos durante el
Pleistoceno. Hasta la fecha solo se reconoce una
especie, X. hodsonae.
|
 |
Este felino medía entre 1.7 a 1.8 metros de
longitud con un cuerpo sumamente musculoso
que probablemente pesaría entre 230 a 400
kilogramos.
Entre las demás especies de felinos dientes
de sable solo Smilodon populator era
de un tamańo mayor. Antes de su
descubrimiento, se pensaba que todos los
felinos dientes de sable caían en dos
categorías. |
Una
incluía a los dientes de sable con largos dientes
caninos superiores y patas cortas y robustas. Los
felinos dientes de cimitarra en cambio tenían
caninos moderadamente alargados y patas largas.
Xenosmilus por su parte poseía tanto patas
robustas y musculosas como caninos cortos y anchos.
Junto a los dos esqueletos originales se hallaron
docenas de huesos de pecaríes. Parece probable que
estos fueran la presa principal de X. hodsonae,
a los que atraparía gracias a su robusta
constitución.
Smilodon.
Lund, 1842.
es
un género extinto de félido dientes de sable de la
subfamilia de los macairodóntinos. El macho más
grande de la especie Smilodon populator pudo
pesar hasta 300 kg, rivalizando con el tigre moderno
por el título de félido más grande de todos los
tiempos.
|
 |
Apareció en América del Norte a finales del
Plioceno y se extinguió en América del Sur
durante el Gran intercambio americano. La
característica más distintiva son sus
enormes caninos, que lo convierten en uno de
los mamíferos prehistóricos más popular.
En la época de transición entre el Plioceno
y el Pleistoceno, los continentes se
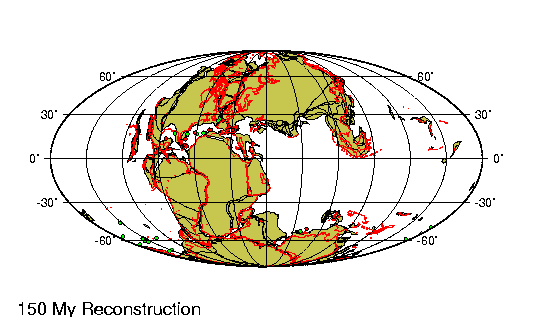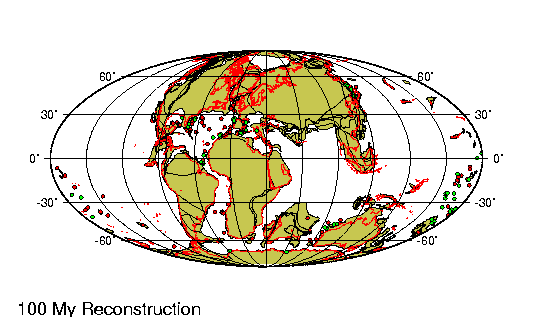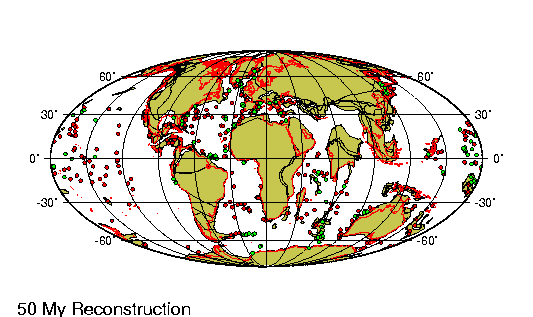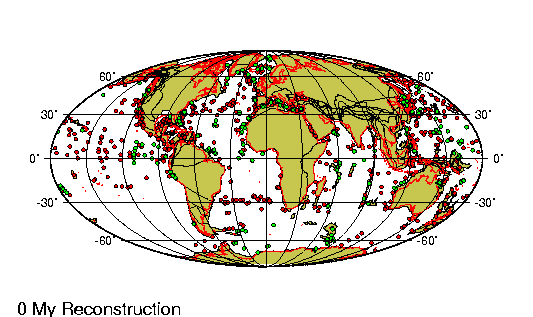
encontraban prácticamente en la misma
posición que tienen actualmente. Las placas
tectónicas sobre las que reposan
probablemente no se han movido más de 100 km
en relación las unas a las otras desde
principios del Pleistoceno. |
A
veces se les denomina «gatos de dientes de sable»
(término impreciso, ya que existieron otros felinos
dientes de sable que no estaban relacionados) o
«tigres de dientes de sable» (también incorrecto
porque tampoco eran tigres). Se extinguieron a raíz
de los cambios climáticos que tuvieron lugar a
finales del Pleistoceno y la modificación de los
ecosistemas que ocasionaron estos cambios, pero es
posible que la llegada de los humanos también
contribuyera a su extinción.
Thylacosmilus.
Riggs,
1933.
Si
hay una prueba validad de convergencia adaptativa,
es Thylacosmilus, el único marsupial del
mundo con dientes de sable. Es un género extinto de
mamíferos marsupiales perteneciente a los
Sparassodonta. Fue un supedepredador tipo dientes de
sable que vivió durante el Mioceno hace
aproximadamente 7 millones de ańos. Los restos de
estos animales han sido hallados en América del Sur.
|
 |
Fue un importante carnívoro marsupial de la
Era Cenozoica, muy parecido al Smilodon
invasor procedente de Laurasia, pero sin
parentesco alguno, ya que este último es un
placentario.
Es otro ejemplo de evolución paralela o
convergencia evolutiva, fenómeno por el cual
dos especies sin parentesco evolutivo y que
nunca habitaron el mismo continente acaban
siendo similares ya que ambas cumplían el
mismo rol en el ecosistema que habitaban. |
Pese a que este animal evolucionó de animales
similares a la zarigüeya en Sudamérica cuando ésta
era un continente-isla, sus características le
hacían casi idéntico a los Machairodontinae (tigres
dientes de sable) que paralelamente evolucionaban en
Norteamérica.
Es
muy posible que sus víctimas fueran los grandes
mamíferos notoungulados como Trigodon
(semejante al rinoceronte), los cuales mataban con
un simple mordisco en el cuello, interrumpiendo el
flujo sanguíneo al cerebro.
Bibliografía sugerida.
Garrido, G. y Arribas, A. (2008). «Generalidades
sobre los carnívoros del Villafranquiense superior
en relación con el registro fósil de Fonelas P-1».
Cuadernos del Museo Geológico y Minero de Espańa
(10): 85–146.
J.
J. Flynn and H. Galiano.
1982. Phylogeny of early Tertiary
Carnivora, with a description of a new species of
Protictis from the middle Eocene of northwestern
Wyoming. American Museum Novitates 2725:1-640
L. D. Martin J. P. Babiarz, V. L.
Naples, J. Hearst: Three Ways To Be a Saber-Toothed
Cat. In: Naturwissenschaften. 87, 1, 2000, , S.
41–44,
Legendre, S.; Roth, C. (1988).
«Correlation of carnassial tooth size and body
weight in recent carnivores (Mammalia)». Historical
Biology 1 (1): 85–98.
H. N. Bryant. 1991. Phylogenetic
relationships and systematics of the Nimravidae (Carnivora).
Journal of Mammalogy
Magnussen Saffer, Mariano.2015. La
Era de Hielo; desde la gran pantalla a la sala del
museo. Paleo Revista Argentina de
Paleontología. Boletín Paleontológico. Ańo XIII.
127: 16-28.
Rincón, A., Prevosti, F., & Parra, G. (2011).
New saber-toothed cat records (Felidae:
Machairodontinae) for the Pleistocene of Venezuela,
and the Great American Biotic Interchange. Journal
of Vertebrate Paleontology, 31 (2), 468-478
Riggs, E.C. 1934: A new marsupial
saber-tooth from the Pliocene of Argentina and its
relationships to other South American predacious
marsupials. Transactions of the American
Philosophical Society 24, 1–32.
Steenhuysen, Jules (2007). «Smilodon
had wimpy bite but teeth to die for». Reuters.
Consultado el 19 de abril de 2008.
Turner, Alan (1997). The Big Cats and
their fossil relatives. New York: Columbia
University Press. p. 60.
Werdelin, Lars & Lewis, Margaret E.
(2001), «A revision of the genus Dinofelis (Mammalia,
Felidae)», Zool. J. Linn. Soc. 132 (2): 147–258,
Los Peces Celacantimorfos o
Celecantos,
nadadores arcaicos
o fósiles vivientes.
Por
Mariano Magnussen Saffer. Director de Grupo
Paleo y Presidente de la Asociación de Amigos del
Museo Municipal Punta Hermengo.
marianomagnussen@yahoo.com.ar. Paleo,
Revista Argentina de Paleontología. Boletín Paleontológico.
Ańo 9. 63: 16-18.
No
todos los días aparece un fósil viviente en las
redes de un pescador. Pero así sucedió en 1938,
cuando Marjorie Courtenay-Latimer, conservadora de
un museo sudafricano, observó una insólita criatura
con gruesas escamas, extrańas aletas y un lóbulo de
más en la aleta caudal, entre una captura de peces
corrientes. Aunque no lo supo de inmediato, había
redescubierto el celacanto, que se creía extinguido
desde finales del cretácico pero que en realidad
sobrevivió a muchos de sus coetáneos prehistóricos y
había seguido habitando las profundidades del
océano, imperturbable –e invisible– durante
milenios.
Los
celacantimorfos (Coelacanthimorpha) o celacantos son
peces de aletas lobuladas (Sarcopterigios) que se
creían extintos (se trata también por tanto de un
relicto) desde el período Cretácico hasta que, en
1938, un ejemplar vivo fue capturado en la costa
oriental de Sudáfrica. Y otra especie que se
localizó en Célebes (Indonesia) en 1998. Junto con
los peces pulmonados, son los seres vivos marinos
más cercanos de los vertebrados terrestres.
Aparecieron en el período Devónico, hace 400
millones de ańos, aunque la mayor cantidad de restos
fosilizados pertenecen al período Carbonífero, es
decir, hace 350 millones de ańos.
 |
El 22 de diciembre de 1938 se descubrió el
primer ejemplar contemporáneo de este grupo
fósil, 80 millones de ańos después del
último registro fósil en que aparecían
ejemplares de este grupo. Fue capturado por
pescadores a unos 60 metros de profundidad
ante la desembocadura del río Chalumna, en
el Sur de África. Medía 1,5 metros de
longitud y pesaba unos 50 kg. Fue
desembarcado en el puerto de East London, en
la República de Sudáfrica. |
El
ejemplar fue analizado por J. L. B. Smith, de la Rhodes
University de Grahamstown (Sudáfrica), que concluyó que
se trataba de un ejemplar perteneciente al grupo de los
celacantos. Lo llamó Latimeria chalumnae,
haciendo referencia al río Chalumna, y referenciando en
el nombre genérico a la conservadora del museo de East
London, Marjorie Courtenay-Latimer, que le envió el
ejemplar junto a una serie de dibujos al darse cuenta de
la rareza del animal. Los dibujos se convirtieron en
algo fundamental, al conservarse únicamente el esqueleto
al llegar a manos de Smith.
Smith
puso carteles en varios idiomas por las costas del
Sudeste africano para intentar conseguir otros
ejemplares. En 1952, un capitán mercante obtuvo un
ejemplar capturado por un pescador en las islas Comores,
situadas en el Océano Índico, entre Madagascar y
Mozambique. Estas quedan a 2500 km de Sudáfrica, y no
había vuelos regulares. Smith se desplazó hasta allí con
un avión de la fuerza aérea sudafricana, tras convencer
de la importancia científica del hecho al primer
ministro de Sudáfrica. Gracias a esta rapidez en el
transporte, en este segundo ejemplar ya se pudo realizar
un análisis interno.
En 1987
se tomaron las primeras imágenes submarinas del
Celacanto en su medio natural, mediante un sumergible.
Fue realizada por M. N. Brenton (del instituto J. L. B.
Smith de ictiología) y Hans Fricke, y patrocinada por la
National Geographic Society, y por la revista alemana
Geo. En 1998 Mark V. Erdmann, biólogo de la Universidad
de Berkeley encontró Celacantos en las Islas Célebes a
casi 10.000 kilómetros de distancia de las Islas
Comores. Esto demostró que la región donde habitan es
muy amplia. Tras este descubrimiento, se han descubierto
otras poblaciones a lo largo del Océano Índico y mares
interiores.
 |
En la actualidad, se han capturado numerosos
ejemplares en las zonas próximas a las dos islas
del archipiélago de las Comores, así como en las
Célebes (Indonesia), Kenya, Tanzania,
Mozambique, Madagascar, Islas Célebes y en el St.
Lucia Wetland Park (Sudáfrica). Incluso se han
citado casos de ejemplares diferentes, algunos
con bioluminiscencia. El problema del estudio de
estos ejemplares estaba en que reventaban a
menudo debido a las diferencias de presión, y
las muestras eran muy defectuosas. |
En la
actualidad, se han capturado numerosos ejemplares en las
zonas próximas a las dos islas del archipiélago de las
Comores, así como en las Célebes (Indonesia), Kenya,
Tanzania, Mozambique, Madagascar, Islas Célebes y en el
St. Lucia Wetland Park (Sudáfrica). Incluso se han
citado casos de ejemplares diferentes, algunos con
bioluminiscencia. El problema del estudio de estos
ejemplares estaba en que reventaban a menudo debido a
las diferencias de presión, y las muestras eran muy
defectuosas.
El
celacanto de Comores mide entre 1,5 y 1,8 metros y llega
a pesar entre 65 y 98 kilogramos. Presenta una
coloración negra, un poco azulada, con manchas blancas y
ligeramente rosas. Tiene una complexión de depredador al
acecho, ya que no está adaptado a la velocidad. A pesar
de ser un pez sarcopterigio, con una base muscular de
las aletas, no se arrastra por el fondo marino, sino que
nada lentamente. Su cola es trilobulada, una
característica insólita. Sus escamas son duras, grandes
y gruesas. Posee una segunda aleta dorsal cercana a la
cola. Su cabeza es grande, con una boca ancha y ojos
grandes y negros. El cráneo posee bisagras. La vejiga
natatoria no es funcional y está llena de grasa. Es un
pez que pare vivas a sus crías, tras un largo período de
gestación, aunque su reproducción sigue siendo un
misterio. Las hembras son más grandes que los machos
No se
conoce con exactitud su comportamiento reproductivo. Se
cree que la madurez sexual no ocurre antes de los 20
ańos. La forma de reproducción es ovovivípara, con
fecundación interna, y huevos que llegan a medir 10 cm
de largo y pesar hasta 300 g. El período de gestación es
de alrededor de 13 meses tras el cual la hembra da a luz
entre 5 y 25 crías bastante desarrolladas, capaces de
sobrevivir, sobre las cuales no se realiza ningún tipo
de cuidado parental.
Son
peces depredadores. Durante el día habitan en cuevas
situadas en zonas profundas (de 150 a 300 m), subiendo
por las noches a la superficie, donde se alimentan de
peces de arrecife.
Bibliografía sugerida.
Jewett, Susan L., "On the Trail of the Coelacanth, a
Living Fossil", The Washington Post, 1998-11-11,
Retrieved on 2007-06-19.
pages 63-82, Weinberg, Samantha. 2006. A Fish Caught in
Time: the Search for the Coelacanth. HarperCollins
Publishers, New York, NY.
Origen y diversidad de los mamíferos
durante el Terciario, desde el Periodo Paleoceno al
Periodo Pleistoceno.
Tomado por; Magnussen Saffer,
Mariano (2010).
Origen y diversidad de los mamíferos durante el Terciario,
desde el Periodo Paleoceno al Periodo Pleistoceno. Paleo, Boletín
Paleontológico. Ańo 8. 45: 17-20.
marianomagnussen@yahoo.com.ar
Los mamíferos (Mammalia) son
una clase de vertebrados amniotas homeotermos (de
"sangre caliente"), con pelo y glándulas mamarias
productoras de leche con la que alimentan a las crías.
La mayoría son vivíparos (con la notable excepción de
los monotremas: ornitorrinco y equidnas). Se trata de un
taxón monofilético; es decir, todos descienden de un
antepasado común que se remonta a finales del Triásico,
hace más de 200 millones de ańos. Pertenecen al clado
sinápsidos, que incluye también numerosos "reptiles"
emparentados con los mamíferos, como los pelicosaurios y
los cinodontos. Se conocen unas 5.416 especies actuales,
de las cuales 5 son monotremas, 272 son marsupiales y el
resto, 5.139 son placentarios. La ciencia que estudia
los mamíferos se denomina teriología, mamiferología ó
mastozoología.
|
 |
Se desconoce con
exactitud, cuándo y dónde aparecieron los
monotremas en la Tierra. La mayor parte de los
fósiles han sido encontrados en Australia, si
bien recientemente han sido descubiertos en
Argentina dientes fósiles de un ornitorrinco
perteneciente al género Monotrematum
sudamericanum lo que sugiere que se
distribuían por las regiones más meridionales de
Gondwana. El fósil más antiguo del que se
dispone de un monotrema data de más de 100
millones de ańos. Se trata de una porción de la
mandíbula de un Steropodon galmani,
un antepasado del ornitorrinco que debía de ser
del tamańo de un gato. |
Los primeros metaterios eran
parecidos a las actuales zarigüeyas (Didelphimorphia) y
surgieron sobre las tierras de la actual Norteamérica
hace entre 80 y 100 millones de ańos. Desde allí se
extendieron por todos los continentes, pero fue al sur
donde proliferaron sin dificultad y a través de la unión
de estas tierras con el continente Australiano, lo
colonizaron poco antes de que ambos se separaran
definitivamente, quedando aislados del resto de los
mamíferos. Por su parte, los primeros euterios eran
similares a las actuales musarańas (Soricomorpha). Se
desconoce su origen cierto y durante algunos millones de
ańos tuvieron que convivir con los dinosaurios,
evolucionando lentamente hasta que éstos acabaron por
desaparecer y los mamíferos tuvieron que evolucionar
para colonizar la totalidad de la tierra, prácticamente
a su disposición.
El Cretáceo termina con la
desaparición de los dinosaurios hace alrededor de 65
millones de ańos, el clima frío dominante va
suavizándose y la supremacía de los primitivos
mamíferos, más evolucionados que los reptiles, con
capacidad para regular la temperatura corporal, un
sistema nervioso central más desarrollado y un organismo
preparado para adaptarse fácilmente a las condiciones
ambientales hace que éstos se diversifiquen y
evoluciones rápidamente para aprovechar los recursos de
un planeta que los dinosaurios al desaparecer han dejado
virtualmente a su disposición.
El primer primate conocido,
el Purgatorius, aparece en Norteamérica a
finales de este periodo y era parecido a las actuales
tupayas (Macroscelidea).
|
 |
De época similar es
el Meniscotherium¸ representante
de la extinta orden Condylarthra, de la que
surgieron ungulados y carnívoros. Así durante el
Terciario, una vez que metaterios y euterios han
diferenciado sus características, se produce la
explosión de los primeros, debido
fundamentalmente al mayor grado de desarrollo de
los recién nacidos y de independencia de las
crías, lo que aumenta las posibilidades de
supervivencia de la especie. |
Es en el continente
australiano donde, el aislamiento y la ausencia de
competidores placentados, hizo que alcanzaran su mayor
grado de desarrollo y evolución. Durante el primer
periodo de la era Terciaria, el Paleoceno, los mamíferos
mantienen las mismas características del Cretáceo,
apareciendo los desdentados en América del Sur de los
que derivarán las órdenes Pilosa y Cingulata.
No obstante, al comienzo del
Eoceno, siguiente periodo de la era Terciaria, ya habían
aparecido todas las órdenes actuales. Los creodóntidos,
carnívoros del tamańo entre un perro y un león, sentaron
las bases para la posterior evolución de los carnívoros.
También datan de esta época los fósiles de
Zeuglodon, un antepasado de las ballenas que
llegaba a medir veinte metros, y los de uintaterios y
titanoterios, parecidos a los actuales rinocerontes, y
los de calicoteros, precursores de los ungulados.
También surge el Eohippus que es
considerado el precursor de todos los équidos actuales.
El Mesohippus, del tamańo de un cerdo y
sin probóscide, es considerado predecesor de los
modernos elefantes y habitaba en las tierras del actual
Egipto. El Icaronycteris es el más antiguo
murciélago conocido y sus restos han sido hallados en
Norteamérica. Pero fueron los primates los que
conocieron mayor grado de evolución durante esta época,
adquiriendo hábitos arborícolas, y aunque sin el
desarrollo de los actuales simios, si podrían compararse
a los lémures y társidos actuales.
Durante el siguiente
periodo, el Oligoceno, se expandieron los animales
surgidos durante el Eoceno. Los elefantes aumentaron su
tamańo y se dispersaron. El Eohippus dio
paso al Mesohippus, de mayor tamańo. Los
roedores y lagomorfos evolucionaron hasta alcanzar
estados similares a las actuales ratas, conejos y
liebres. Evolucionaba el Hyaenodon,
precursor de las actuales hienas y surgieron el primer
félido, el Dinictis, y el primer cánido,
el Pseudocynodictis. De este periodo son
los fósiles del mamífero terrestre más grande conocido,
el Indricotherium, encontrados en Asia,
con una envergadura superior a los cinco metros y una
longitud de ocho. Los primitivos Condylarthra fueron
dejando paso a los actuales artiodáctilos, siendo el más
antiguo conocido el Oreodontos, de hábitos acuáticos y
poblador de Norteamérica. Los antracoterios, o
hipopótamos primitivos se expandieron sustituyendo a los
rinocerontes acuáticos. Los dos últimos periodos del
Terciario conocieron el mayor florecimiento de los
mamíferos debido fundamentalmente a un enfriamiento del
clima que permitió el surgimiento de grandes extensiones
de pradera capaces de sustentar a gran número de
herbívoros que servían a su vez de sustento a otro
nutrido grupo de carnívoros.
|
 |
Durante el Mioceno, el
intercambio de especies entre Eurasia y África
debido al encuentro de ambas placas
continentales hizo posible que los elefantes
llegaran hasta Norteamérica, mientras que
félidos, cánidos y ungulados vegetarianos
invadieron África. En este periodo La Tierra
adquirió el aspecto actual y la diversidad
animal conoció un esplendor no repetido en
épocas anteriores ni posteriores. Numerosos
ungulados adquirieron sus formas actuales, y
existían además otros con las más variadas y
extrańas cornamentas, como el
Synthetoceras norteamericano con un solo
cuerno en forma de Y o el Syndyoceras
en forma de V. |
Los elefantes eran muy
numerosos y existía una considerable diversidad de
especies, el caballo continúa su evolución a través del
Merychippus y el Pliohippus.
Los más espectaculares animales de la época eran sin
duda los tigres dientes de sable de los géneros
Machairodus y Smilodon y como
otros carnívoros, fueron evolucionando a lo largo del
Mioceno y el Plioceno. También en el Mioceno, los
primates sufrieron una fuerte evolución, además de los
hábitos arbóreos, colonizaron sabanas y estepas y dieron
lugar a la aparición de los antepasados de los homínidos,
considerándose al Ramapithecus el
predecesor de todos ellos.
Con la llegada del
Pleistoceno, comienza la era Cuaternaria, caracterizada
por la aparición de los homínidos que coincide con una
recesión drástica de la diversidad de especies de
mamíferos. Durante este periodo la evolución cambió de
rumbo y en lugar de proceder a incrementar la diversidad
de especies, éstas fueron reduciéndose en pro de una
mayor capacidad de adaptabilidad a los rápidos cambios
climáticos que estaban sucediéndose. Desaparecieron así
numerosas especies y se perfeccionaron las existentes
hasta llegar a la aparición del hombre (Homo
sapiens) actual, causante directo de la rápida
desaparición de numerosas especies animales sean o no
competidoras de la suya.
Bibliografía sugerida.
Brands, S.J. (comp.) (2005)
Systema Naturae 2000. The Taxonomicon Universal
Taxonomic Services, Amsterdam, Holanda. Acceso: 31 de
octubre de 2007.
MacDonald, D. (2001).
The Encyclopedia of Mammals. New York.
Facts on File.
Martin, R.E., Pine, R.H. and
DeBlase A.F. (2001). A Manual of Mammalogy. McGraw-Hill.
San Francisco.
Novak, R.M. (1999). Walker’s
Mammals of the World. John Hopkins University Press.
Baltimore.
Vaughan, T., Ryan, J., and
Czaplewski, N. (1999). Mammalogy. Saunders College
Publishing. Philadelphia.
Grzimek, B., Schlager, N. y
Olendorf, D. (2003). Grzimek's Animal Life Encyclopedia.
Thomson Gale.Detroit.
Pough, F. H., C. M. Janis, and
J. B. Heiser. Vertebrate Life. 6th ed. Upper Saddle
River, NJ: Prentice Hall, 2001.
Soriano; Pascual J. y Ochoa G,
José. 1997: “Lista actualizada de los mamiferos de
Venezuela”. En: La Marca, E. (Editor): Vertebrados
actuales y fósiles de Venezuela. Serie Catalogo
Zoológico de Venezuela. Vol. 1. Museo de Ciencia y
Tecnología de Mérida. Mérida – Venezuela. Pp:205-227.ISBN
980-221-877-4
Young, J. Z. 1977. La vida de
los vertebrados. Editorial Omega, Barcelona, 660 pp.
Agualinda, Colombia. Un tesoro Paleontológico |
|
en peligro de desaparecer.
|
|
agualinda colombia fosiles
paleontologia agualinda colombia fosiles paleontologia agualinda
colombia fosiles paleontologia
|
|
Por
Jorge Villamizar.
diovenganza@gmail.com.
|
|
Articulo completo publicado originalmente en Paleo, Boletín
Paleontológico. Ańo 5. Numero 28: 13 - 15.
|
|
agualinda colombia fosiles paleontologia agualinda colombia
fosiles paleontologia agualinda colombia fosiles paleontologia
|
Quisiera contarles a través de este artículo la importancia de
esta zona para el mundo arqueológico, paleontológico y
ecoturistico, un lugar que se puede considerar como un tesoro
que guarda en su interior joyas y secretos de gran importancia
para el interés científico y cultural de Colombia y el mundo.
Agualinda esta ubicada en el Departamento de Norte de Santander
en el municipio de Los Patios- Colombia, es un lugar con una
belleza única que muy pocos ven en realidad ya que nadie se
atreve a ver mas allá de sus intereses económicos y no ven el
legado cultural que destruyen a diario digo esto porque
Agualinda es una zona rica en arcilla y canteras de piedra
caliza y las empresas que trabajan en la zona no tienen un
control de explotación adecuado y cada vez se expanden mas,
reduciendo las posibilidades de conservación y estudio.
|
 |
Pero no todo es malo existe una persona que se enamoro
de esta tierra así como de su esposa alguien que conoce
su belleza y secretos como ninguno y que muy pocos
cuestionan el conocimiento y la experiencia que posee de
la región, su nombre es Saddy Molina Escalante o el
seńor de los fósiles, una persona que no es de esta
región pero que llego a ella y se quedo y que durante
mas de 40 ańos se ha dedicado a recolectar y estudiar
las diferentes piezas fósiles que conforman el museo que
con esfuerzo y dedicación a creado.
El museo posee
alrededor de 230 piezas de diferentes periodos geológico
entre las que encontramos: |
FÓSILES MARINOS; Ammonites del periodo jurasico,
Bivalvos del periodo cretácico, Gardiolas interruptas
(siluriano superior), Ganoideos (Devoniano),
Amnatos del periodo Cretácico, Cola de pez teleósteos
(Eoceno), Cabeza de pez teleósteo (Eoceno).
FÓSILES SIN IDENTIFICAR; Fragmentos de costillas (pertenecen a
un reptil marino), Diente de reptil sin identificar, Esqueleto
casi completo de un reptil marino de 7 metros sin identificar y
que a un no a sido extraído del lugar de yacimiento.
Por el momento todos se preguntaran que tiene de especial este
lugar ya que los fósiles que les acabo de mencionar se
consiguen en las mayorías de yacimientos del mundo pues les
diré porque este lugar es muy importante para Colombia como
yacimiento paleontológico.
 |
MEGAFAUNA; Superfamilia: Glyptodontoidea, familia
gliptodóntidae genero y especie indeterminada aunque el
material encontrado es pobre (4 placas incompletas) no
obstante, es posible que las placas puedan referirse a
la subfamilia Sclerocalyptinae. Por otra parte es
necesario resaltar la importancia del descubrimiento,
pues constituye el primer registro de gliptodóntido
pliocénico de Colombia.
También existe material recolectado de especies como
Toxodonte de la especie Mixotoxodon larensis,
Odocoileus (orden artiodactyla), también
existen restos de Mastodonte (Haplomastodon
waringi). |
Pero el mas importante de todos los descubrimientos en cuanto a
megafauna es el del Eremotherium (Pseudoeremotherium)
cucutense Porta 1961 según los registros Porta en
el ańo de 1961 describe restos de megaterinos procedentes de
Cúcuta; considera que el material encontrado constituye una
nueva especie y propone la especie como Eremotherium (Pseudoeremotherium)
cucutense ańos después ilustran dientes aislados de
Eremotherium que fueron recolectados en el lugar
denominado las piletas, dentro de la vereda Agualinda, municipio
de Los Patios
También se ha recolectado material lítico de los primeros
humanos que habitaron esta zona (cazadores recolectores)
ECOTURISMO
Agualinda no solo es una tierra rica en yacimiento fósiles
también posee un atractivo muy especial unos colosos que
descansan entre el follaje de la zona ellos son Los Estoraques
formaciones de arcilla que han sido talladas cuidadosamente por
el viento y el agua los cuales contrastan con el paisaje de la
zona cautivando a los visitantes con su tamańo y misterio
 |
Como ven es una tierra con un gran potencial
arqueológico, paleontológico y ecoturistico que lo único
que necesita mayor apoyo en investigación y recursos ya
sean nacionales o internacionales lo importante es
conservar este lugar para la prosperidad.
Actualmente solo se esta trabajando en un proyecto de
grado con los estudiantes de la Corbes, Natalia Masmela,
Yajaira Abreu y su servidor Jorge Villamizar y con la
colaboración de don Saddy Molina Escalante el cual
consiste en creación de un parque arqueológico y
paleontológico en donde se pueda mostrar al visitante
las maravillas de este lugar. |
Pero por el momento solo esta en el papel ya que no existen
recursos ni ayudas de ningún tipo por eso si algún lector esta
interesado o conoce alguna empresa que ayude a la conservación
de estos lugares especiales me escriban a mi correo
jvm49@hotmail.com
se les agradece su colaboración.
|
agualinda
colombia fosiles paleontologia agualinda colombia
fosiles paleontologia agualinda colombia fosiles
paleontologia |
Dinosaurios poiquilotermos u homeotermos. żEran o
no de sangre caliente?
Tomado de;
Magnussen Saffer,
Mariano (2010).
Dinosaurios poiquilotermos u homeotermos. żEran o no de
sangre caliente?. Paleo, Boletín
Paleontológico. Ańo 8. 43: 13-16.
marianomagnussen@yahoo.com.ar
Un animal de
sangre caliente sería aquel que normalmente mantiene su
temperatura corporal casi constante, dentro de estrechos
límites. Y si los dinosaurios están estrechamente emparentados
con las aves (algo de lo que quedan ya muy pocas dudas, en vista
del abrumador número de pruebas que se han ido acumulando), es
posible que su temperatura fuera realmente alta, ya que las aves
son los vertebrados que tienen ajustado su termostato interno a
mayor temperatura, en muchos casos a más de 42 ş C
El nombre
correcto de estos seres es poiquilotermos y abarca a todos los
reptiles. Una lagartija toma el sol para calentar su cuerpo y
así poder moverse más rápido, lo que significa que en un día
frío será más lenta. Lo contrario ocurre con los animales de
“sangre caliente” u homeotermos, los cuales mantienen una
temperatura constante en sus cuerpos independientemente de las
condiciones del medio ambiente (tal es el caso de los seres
humanos y todos los mamíferos).
Cuando se
descubrieron los primeros fósiles de dinosaurio, todo el mundo
comprendió que eran reptiles. Y como todos los reptiles actuales
son de “sangre fría”, muchos expertos piensan que ése también
fue el caso de los dinosaurios. Sin embargo, algunos científicos
creen que muchos de ellos tuvieron que ser veloces y ágiles
cazadores de “sangre caliente” e incluso que algunas especies
estaban cubiertas de plumas o de pelo. En la actualidad, los
científicos mantienen un animado debate sobre estas dos teorías
sin llegar a un acuerdo concreto.
|
 |
Ésta y
otras preguntas tratan de ser contestadas por los
paleontólogos, que son las personas encargadas de
estudiar los fósiles. Aunque te parezca divertida, esta
labor puede ser realmente muy difícil, ya que en la
mayoría de los casos los restos de un animal
prehistórico se encuentran diseminados en un terreno muy
extenso (de hecho, los fósiles de esqueletos enteros son
muy raros). Los paleontólogos identifican la especie a
la que pertenecieron los huesos fósiles, los extraen del
terreno, los ordenan como si formaran parte de un
rompecabezas y determinan su antigüedad.
Los
dinosaurios se encuentran entre los animales de mayor
éxito en la historia de nuestro planeta. Dominaron la
tierra durante más de 160 millones de ańos... un período
extraordinariamente largo, sobre todo si consideramos
que los seres humanos apenas existimos desde hace tres
millones de ańos. |
Las tomografías
computerizadas que se realizaron a este ejemplar mostraron un
órgano en la cavidad torácica que parece corresponder (las dudas
son escasas) a un corazón. Se observan 4 cámaras y una única
aorta, como en aves y mamíferos y a diferencia de los reptiles.
Esto sugiere que poseía unos sistemas circulatorio y pulmonar
separados y un metabolismo más rápido que el de los reptiles, lo
que podría permitirle mantener alta su temperatura.
Al parecer la
fama de que los grandes terópodos carnívoros como el
Tyranosaurus, Gigantosaurus, el
Carnotaurus y otros mas pequeńos como el
Velocirraptor (solo por citar algunos) de haber sido
unos
depredadores de sangre fría es infundada, así lo
demuestra un reciente estudio el cual sugiere que su sangre era
caliente, como la de las
aves
y
mamíferos.
Los paleontólogos han debatido durante décadas acerca del
metabolismo de los dinosaurios : żEstos antiguos y pesados
animales utilizaban principalmente en el calor del sol para
regular su temperatura corporal como los reptiles y anfibios de
hoy, o podrían generar su propio calor corporal como los
mamíferos y las aves?.
Ahora, usando
un modelo biomecánico que predice el coste energético de caminar
y correr en función del tamańo de los huesos de las patas de un
animal, los investigadores han demostrado que los mayores
dinosaurios no habrían conseguido tener esos huesos sin un
metabolismo de sangre caliente. De los estudios realizados sobre
animales vivos, podemos calcular la relación entre el diseńo de
las extremidades y la cantidad de músculos que un animal
necesita para soportar su peso corporal.
"El tamańo del
músculo sirve para predecir la cantidad de energía que necesita,
porque a su vez en el músculo, necesita oxígeno. Debido a que
los animales de sangre caliente tienen una capacidad aeróbica
mayor que sus homólogos de sangre fría, la búsqueda de músculos
más grandes y mayores demandas de energía en los dinosaurios
estaría a favor de la hipótesis de que eran
animales
de sangre caliente. De hecho cuando se
analizaron los modelos anatómicos de 14 especies diferentes
dinosaurios, sorprende al encontrar que incluso a paso lento, la
mayoría de los dinosaurios necesitan más energía para
desplazarse que la producida por un metabolismo de sangre fría.
|
 |
Para
sacar conclusiones acerca de dinosaurios extintos hace
millones de ańos basado en un modelo de animales que
existen en la actualidad es necesario hacer algunas
suposiciones. Algunos investigadores reconocen que es
posible dinosaurios tuviesen una
fisiología completamente diferente a
todos los seres vivos de hoy, un metabolismo de sangre
fría que proporcione la energía suficiente para
satisfacer sus necesidades aeróbicas. Esa es una
limitación de este análisis, y tal vez una limitación de
cualquier análisis similar basado en animales de la
actualidad. |
La nueva
investigación concuerda con un estudio previo sobre la anatomía
cardiovascular de los dinosaurios, basado en una tomografía
computarizada de un corazón fósil de 66 millones de ańos. Las
imágenes revelaron un corazón de doble bomba con una sola aorta,
en esencia, el corazón de un mamífero de sangre caliente o un
pájaro, y no un reptil de sangre fría. Sin embargo, otros
estudios anatómicos han llevado a conclusiones diferentes: un
estudio sobre la nariz de los dinosaurios, comprobó que carecía
de unos huesos llamados cornetes, que protegen contra la pérdida
de agua durante la respiración rápida y se encuentran en 99 por
ciento de los animales de sangre caliente.
El ser humano
está dentro de este grupo, ya que como todos sabemos, tenemos
una temperatura cercana a los 37ş C. Nuestro organismo trata de
mantener siempre esta temperatura porque es la óptima para que
trabajen nuestras células, y por tanto para nuestra
supervivencia. En cambio, los animales de
sangre
fría siempre habían sido agrupados como los que
varían su temperatura interna en función de las
condiciones ambientales, dejando a un lado cualquier control
interno. Dentro de este grupo podríamos encontrar los lagartos.
Algunos de estos animales, incapaces de controlar su temperatura
interna, necesitan acumular calor durante las horas de sol para
mantenerlo en las horas más frías.
En un extremo,
los animales con gran control de su temperatura interna. En el
otro, los que son incapaces de realizar cualquier control
interno, por lo que son totalmente dependientes del ambiente
exterior. Lo que era “sangre caliente”, pasó a distribuirse en
tres grandes grupos: Endotermia, Homeotermia y Taquimetabolismo.
Y lo que se entendía como “sangre fría”, pasó a separarse en lo
siguiente: Ectotermia, Poiquilotermia y Bradimetabolismo.
Pues bien,
desde los ańos setenta, Ostrom y Bakker mantienen que los
dinosaurios eran animales de sangre caliente, como los mamíferos
y las aves, iniciando una larga controversia aún no resuelta,
hoy dormida, aunque el hallazgo, este ańo, de dinosaurios
celurosaurios con plumas, a mi modo de ver, la zanja
parcialmente a favor de los dinosaurios de sangre caliente. Al
menos estos lo eran sin lugar a dudas, puesto que el aislamiento
térmico que suponen las plumas sería contraproducente para un
animal de sangre fría, que dependiera del calor exterior para
mantener su temperatura.
Las pruebas
principales a favor de la endotermia de los dinosaurios son
cinco: La estructura interna de los huesos. El hueso es una
estructura activa, en crecimiento y fisiológicamente importante,
puesto que en su médula se fabrican células sanguíneas y el
tejido óseo es un reservorio de calcio movilizable para la
contracción muscular y otros procesos importantes. Los animales
con un alto nivel metabólico y procesos fisiológicos acelerados
exigen mayor rendimiento de los huesos. En los animales
endotermos que conocemos, los vasos que penetran en los huesos
largos son numerosos y densamente agrupados; en los reptiles y
anfibios, son escasos. Los conductos de Havers, donde se da el
intercambio de calcio entre el hueso y la sangre son más
numerosos en los huesos de mamíferos y aves que en los reptiles
y anfibios. Ricqlčs (1969) afirmó que la estructura interna de
los huesos de los dinosaurios se parecía más a la de los
mamíferos y aves que a la de reptiles y anfibios. Aunque sus
apreciaciones son objeto de discusión.
|
 |
Ausencia de anillos de crecimiento en los huesos de los
dinosaurios que vivieron en latitudes elevadas. Esas
latitudes, aunque más cálidas que las actuales, no
dejarían de tener fríos inviernos y grandes variaciones
estacionales de temperatura. En la estación fría, el
hueso de los reptiles y de los anfibios crece más
lentamente, debido a un metabolismo muy ralentizado, que
en la estación cálida, dando lugar a anillos de
crecimiento. Dado que los huesos de los dinosaurios de
esas latitudes no presentan tales anillos, no debían de
ser de sangre fría.
La
visión que se tiene actualmente de los dinosaurios como
criaturas activas, con los miembros situados bajo el
cuerpo en vez de salir horizontalmente del mismo, a
diferencia de lo que sucede con los reptiles actuales.
Esto les asemejaría a los mamíferos corredores en la
postura y en las proporciones de sus extremidades. Una
postura y una locomoción de este tipo no podrían
mantenerlos unos animales ectotermos, de bajo
metabolismo, del tamańo de la mayoría de los
dinosaurios. |
En nuestra era,
ningún gran animal ectotermo vive en latitudes elevadas. El
tiempo que necesitarían para recuperar su temperatura óptima
sería larguísimo (sin contar que cada día tiene su noche). Sin
embargo, hay fósiles de grandes dinosaurios en lo que fue en su
tiempo el círculo ártico, con su medio ańo de ausencia de sol.
Se ha aducido que emigrarían. Pero la emigración supondría más
de 800 km. żPodría resistirlo un animal de sangre fría?
Nosotros tal
vez podríamos darnos por satisfechos con todas estas pruebas y
admitir ya que los dinosaurios eran de sangre caliente, pero los
científicos son tremendamente meticulosos, puntillosos y
desconfiados (los que no han presentado estas características
han metido la pata en incontables ocasiones) y seguirán buscando
más pruebas. Además, quedan aún muchas preguntas pendientes que
los mantendrán entretenidos presumiblemente durante décadas:
żCuál era su temperatura corporal y su rango de variación? żDe
qué medios se valían para absorber y disipar calor? żRegulaban
su temperatura de forma diferente los dinosaurios grandes y los
pequeńos?. El debate sigue abierto pero este parece volcarse
para el lado de que los dinosaurios de gran tamańo tenían sangre
caliente en sus venas.
Bibliografía
sugerida
Andrés, J.A. y
Pérez-Lorente, F. (2005): La declaración de los yacimientos de
icnitas de dinosaurio de la Península Ibérica como patrimonio
mundial (IDPI). XXI Jornadas de la Sociedad Espańola de
Paleontología. Gestión e Investigación de la Paleontología en el
XXI. Sevilla 4-8 de octubre del 2005.
Bravo, A.M.,
Vila, B., Galobart, A. y Oms, O. (2005): Restos de huevos de
dinosaurio en el Cretácico superior del sinclinal de Vallcebre (Berguedŕ,
provincia de Barcelona). Revista Espańola de Paleontología,
Número extraordinario, 10: 49-57.
Le Loeuff, J. y
Martínez, A. (1997): Afloramiento de icnitas de Titanosauridae
en la zona de Fumanya (Maastrichtiense, Pirineo oriental):
estudio preliminar. Geogaceta, 21: 151-153.
Sanz, J.L.
(1999): Los dinosaurios voladores. Historia evolutiva de las
aves primitivas. Ediciones Libertarias/Prodhufi, S.A. Mundo
Vivo. 239 págs. Sanz, J.L. (1999): Mitología de los dinosaurios.
Editorial Taurus. Pensamiento. 206 págs.
Sanz, J.L.
(2007): Cazadores de dragones. Historia del descubrimiento e
investigación de los dinosaurios. Editorial Ariel. 420 págs.
Sanz, J.L. y Buscalioni, A.D. (Coords.) (1992): Los dinosaurios
y su entorno biótico. Ayuntamiento de Cuenca, Instituto "Juan de
Valdés". Actas académicas, 4. 397 págs.
Sanz, J.L.,
Buscalioni, A.D., Moratalla, J.J., Francés, V. y Antón, M.
(1990): Los reptiles mesozoicos del registro espańol. C.S.I.C.
Museo Nacional de Ciencias Naturales. Monografías, 2. 79 págs.
Zaera, Julio
Loras. Reptiles de sangre caliente? http://fortanete.cjb.net/_cms_repcal.html
|
agualinda
colombia fosiles paleontologia agualinda colombia
fosiles paleontologia agualinda colombia fosiles
paleontologia |
Algunas consideraciones sobre las icnitas como
testimonio fósil.
Tomado de;
Magnussen Saffer,
Mariano (2012). Algunas
consideraciones sobre las icnitas como testimonio fósil. Paleo,
Revista Argentina de Paleontología. Boletín Paleontológico.
Ańo 10. 67: 30-33.
marianomagnussen@yahoo.com.ar
Una icnita es
cualquier huella o seńal producto de la actividad biológica,
especialmente pistas, rastros, pisadas, madrigueras, orificios,
perforaciones, seńales de mordeduras, etc. Se clasifican en
icnotaxones. La disciplina que estudia las icnitas se denomina
Icnología (Paleoicnología para el caso particular de las
fosilizadas).
Las
icnitas
pueden ser las marcas de desplazamiento, como los rastros
continuos dejados por el avance de un crustáceo o gusano, o las
marcas de las pisadas de un dinosaurio. Y aunque generalmente no
se sepa qué especie fue la que produjó la huella, si es posible
determinar el grupo de dinosaurios al que pertenece, e incluso,
tras un estudio detallado, obtener información sobre el tamańo,
comportamiento y ecología del animal que la produjo,
especialmente si se encuentran formando un rastro.
Las seńales
pueden ser de origen biogénico al igual que los restos fósiles,
pero también pueden ser generadas por los restos, es decir, por
el organismo una vez muerto y, por tanto, no son de origen
biogénico sino que tendrían un origen tafogénico. En otras
ocasiones las huellas son producidas por los organismos pero
condicionadas por elementos externos (condiciones climáticas).
Por ejemplo, las estructuras circulares que forman plantas sobre
sedimentos blandos por efecto del viento o marcas dejadas por la
lluvia sobre ceniza volcánica.
Las primeras
icnitas del mundo fueron encontradas en 1802 en Massachusetts,
en una graja a 15 millas al norte de Connecticut (en South
Hadley), por un chico llamado Pliny Moody; encontró un bloque de
piedra con cinco huellas tridáctilas que su familia usó como
ornamento durante algún tiempo. Más gente de la zona encontró
huellas similares, que fueron interpretadas como las de un
pájaro gigante antiguo, algunas de ellas similares a las de un
pavo. Los ciudadanos de Greenfield, en 1835, decidieron colocar
estas piedras en el pavimento de la ciudad, y esto llamó la
atención del Dr.
Edward
Hichtcock, un sacerdote profesor del Amherst
College entre 1825 y 1864, que comenzó a estudiar las huellas y
dedujó, observando su variedad y analizando miles de ellas, que
pertenecían a diversos grupos de organismos, que erroneamente
atribuyó a aves; su trabajo, hoy un clásico, describe con
precisión y minucioso detalle las huellas encontradas, y fue
publicado en 1858 bajo el título
Ichnology of New England.
 |
En
Europa fue
Henry Duncan (1774-1846) el pionero; era
un clérigo, filósofo, escritor, arqueólogo y otras
varias más; en 1824, mientras ejercía como sacerdote en
Ruthwell (Escocia), le presentaron un bloque de arenisca
roja procedente de Corncockle Muir en Annandale que
tenía varias huellas. Sabiendo que eran las huellas de
un animal, escribió al reverendo William Buckland,
geólogo de la Universidad de Oxford, para pedirle su
opinión, y éste le urgió para publicar su
descubrimiento, lo que tuvo lugar en 1831, siendo ésta
la primera publicación de la evidencia fósil de huellas
de dinosaurio. |
Pero la
personalidad más importante en paleoicnología ha sido
Roland T.
Bird, que, a pesar de no tener una formación en
paleontología, le encantaba buscar fósiles en el oeste americano
entre 1930 y 1950. Fue un importante investigador en Howe Quarry,
Wyoming, el principal yacimiento de huesos de dinosaurios del
mundo, pero es mundialmente conocido por haber descubierto y
estudiado los rastros de dinosaurios del Cretácico en Glen Rose,
Texas, que son los mayores encontrados en EEUU. Sus estudios han
puesto de manifiesto comportamientos en los dinosaurios hasta
entonces no conocidos, entre ellos el gregarismo
Los
icnofósiles se caracterizan por tener una distribución temporal
larga, lo que genera seńales de morfología más sencilla. Hay un
rango de facies restringido, lo que permite la identificación de
facies análogas de intervalos temporales no iguales, gracias a
esto se pueden realizar caracterizaciones paleoambientales. No
sufren procesos de transporte, aunque puede haber desplazamiento
de las seńales si se produce una mineralización temprana. Los
icnofósiles son casi la única fuente de información sobre los
organismos de cuerpo blando.
 |
Se
clasifican
según el grado de consolidación del sustrato: blando,
viscoso, firme y endurecido. Un sustrato blando laminado
adquiere heterogeneidad por perturbaciones, llegando a
una homogeneidad total si éstas son muy intensas. En un
sustrato blando se pueden encontrar dos tipos de
estructuras: Texturas de bioturbación. Reestructuración
de las partículas del sedimento blando, por ejemplo la
acción de determinados gusanos que acumulan partículas
gruesas a determinada profundidad (no se ven los tubos).
Se da una estratificación biogénica. Estructuras de
bioturbación. Generan seńales de cuerpos con morfología
y estructura reconocibles.
Si
el sustrato está más consolidado las cavidades
permanecen estables aunque sean muy verticales y, se ven
las "excavaciones" ya que los organismos han de
perforar, dejando límites erosivos. Se habla de
excavación (burrow, galería) en sustrato firme y, de
perforación (boring) en sustrato endurecido. En un
sustrato firme se produce una reestructuración de las
partículas. |
La
clasificación estratinómica se realiza en función de la posición
de las huellas en el sustrato. Epirrelieve: estructura biogénica
en el techo de la capa o del estrato. Hiporrelieve: estructuras
biogénicas que se observan el muro de la capa o del estrato.
Ambos pueden ser convexos o cóncavos. Endorrelieves: estructuras
biogénicas en el interior del sustrato.
Clasificación
etológica basada en el tipo de actividad reflejada en la
estructura biogénica. Cubichnia: seńales de reposo. Depresiones
de contorno cerrado y poco profundas realizadas por animales que
se han apoyado y parado sobre un sustrato no endurecido.
Generalmente el contorno de la depresión reproduce el contorno
ventrolateral del cuerpo del animal productor o de la parte del
cuerpo usada para apoyarse. Ejemplos: Rusophycus, huella de
contorno cerrado de un trilobites, huellas de estrella de mar.
 |
Domichnia:
cavidades de habitación o de morada. Son estructuras de
bioturbación o perforación usadas de forma más o menos
permanente por el organismo, generalmente suspensívoro
aunque también puede tener otra alimentación. Tubos de
gusanos, en forma de U, verticales o con una porción
distal horizontal, con doble abertura o con una sola
abertura y verticales como en el grupo de los Skolitos.
Morfologías talasinoides: En todos los casos el sustrato es más o menos firme para evitar el
colapso de la cavidad, pero la morfología depende de los
diversos grados de consolidación del sustrato. Así,
Ophiomorpha se da en sustratos más blandos o inestables
(taludes con pendiente) y Spongeliomorpha se da en
sustratos menos blandos. En los límites de la cavidad se
acumulan mucosidades que producen una cementación
temprana. Ejemplo:Callianassa, estrecha la boca
del tubo para estabilizar la cavidad. |
Si se produce aporte de sedimentos alarga el tubo o la zona estrecha de
éste. Si se produce erosión generalmente cierra la cavidad y
genera otra próxima y más consistente.
Fodinichnia:
estructuras de bioturbación realizadas en el interior del
sedimento, inicialmente como huellas de alimentación, pero
también como cavidades de habitación. Se trata de estructuras de
bioturbación con patrones geométricos densos producidos por
organismos endobentónicos que se alimentan del y dentro del
sedimento y, de este modo, quedan protegidos de los depredadores
mientras se alimentan. El ejemplo más común es el de los
Chondrites, en el que el organismo ocupa un tubo principal del
que va haciendo nuevas aberturas dispuestas paralelamente a la
laminación. Rara vez pasan dos veces por el mismo sitio o por
donde han pasado antes otros.
Pascichnia:
pistas de alimentación. Son seńales de alimentación en la
superficie del sedimento que corresponden a huellas o surcos
discontinuos, a menudo simples y muy próximos entre sí. Rara vez
están superpuestos y suelen presentar patrones geométricos
densos, lo que refleja un máximo aprovechamiento de los
nutrientes distribuidos en la superficie del sedimento.
 |
Suelen ser generadas por organismos limnófagos o
micrófagos. Ejemplos:Helminthoida, huella muy
densa; y,Cosmorhaphe, de trayecto irregular y
menos denso.
Repichnia:
restos o seńales de locomoción más o menos continuas.
Son huellas dejadas por organismos que se desplazan
sobre el sustrato. Son surcos o huellas horizontales,
alineadas o de distribución sinuosa que, pueden estar
superpuestas, divididas o ramificadas. En el caso de los
Trilobites se distinguen dos tipos de huellas, Rusoficus
(reposo) y Cruziana (movimiento). LaAmbulichnia es un
caso de Repichnia discontínua. |
Agrichnia:
pistas de alimentación complejas. Referencia a cuidado y cultivo
de organismos. Suelen darse en zonas de máxima batimetría donde
los nutrientes son escasos y se recurre a "plantaciones" de
bacterias y hongos para la bioquimiosíntesis de materia orgánica
(Paleodictyon). Típicas estructuras de bioerosión que se
producen sobre un sustrato duro.
Praedichnia:
estructuras de predación. Perforación debida a un gasterópodo en
el Mioceno.
Fugichnia:
estructuras de escape. Se trata de trazas que son el resultado
de una rápida y brusca migración del productor en dirección
vertical y oblicua a la estratificación, a través del sedimento
y como consecuencia de una sedimentación rápida, una erosión o
el escape frente a un depredador. Esto suponen un cambio brusco
de medio.
Lecturas
sugeridas
Bayon, C. Y Politis, G., 1996, "Estado actual de las
investigaciones en el sitio Monte Hermoso 1 (Prov. de Buenos
Aires)", en Arqueología (Instituto de Ciencias Antropológicas,
Facultad de Filosofia y Letras, Universidad de Buenos Aires),
6:83-116.
Bromley, R. G. 1990. Trace fossils, biology and taphonomy.
Unwin Hyman, London; 280 pags.
Lińán, E. y
Gámez Vintaned, J. A. 1996. Introducción a la paleoicnología de
invertebrados. Zubía, 14, 65-77, 4 fig. Logrońo. ISSN:
0213-4306. [Artículo basado en la conferencia leída en Enciso
(La Rioja) en 1993.]
Gámez Vintaned,
J. A. y Lińán, E. 1996. Revisión de la terminología icnológica
en espańol. Revista Espańola de Paleontología, 11 (2), 155-176,
3 fig., 1 ap. Madrid. ISSN: 0213-6937.
Politis, G.,
1989, "żQuién mató al megaterio ?", en Ciencia Hoy; 1,2: 26-35.
 Ver
mas Artículos de Divulgación Paleontológica Internacional
Ver
mas Artículos de Divulgación Paleontológica Internacional |