El largo origen de los
camélidos.
Por Mariano
Magnussen Saffer. Grupo Paleo. Museo Municipal Punta
Hermengo de Miramar. Fundación Argentavis.
marianomagnussen@yahoo.com.ar. Tomado de; Magnussen Saffer,
Mariano. 2016. El largo origen de
los camélidos. Paleo Revista Argentina de
Paleontología. Boletín Paleontológico. Año XIV. 148: 22-26.
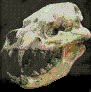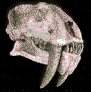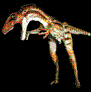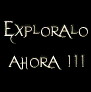
Los
camélidos son estrictamente herbívoros, con largos y
delgados cuellos y piernas prolongadas. Se diferencian de
los rumiantes porque su dentición muestra rastros de
incisivos centrales vestigiales en el maxilar superior, y la
presencia de dientes caninos verdaderos, separados de los
premolares por un espacio llamado diastema, tanto en el
maxilar como en la mandíbula.
Todos los
camélidos, tanto fósiles como actuales, caminan de una
manera particular debido a su sistema de locomoción; en
marcha, las dos extremidades del mismo lado se mueven
simultáneamente, diferentes de los caballos por ejemplo que
tienen un galope intercalado. Los camélidos no tienen
pezuñas; a cambio tienen dos dedos con uñas en cada pie y
almohadillas de plantar, únicos tilópodos.
 |
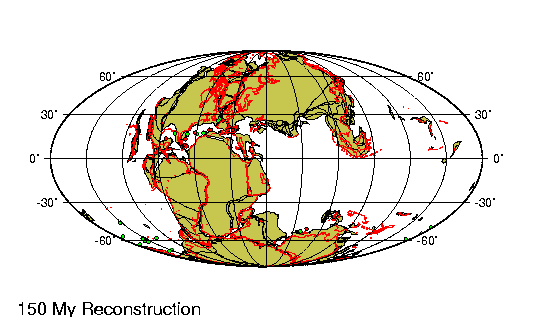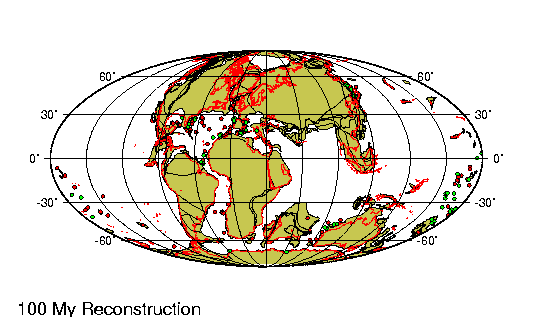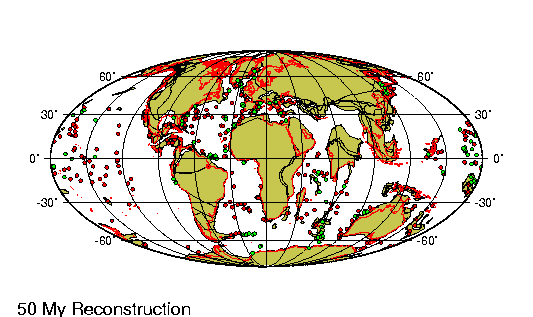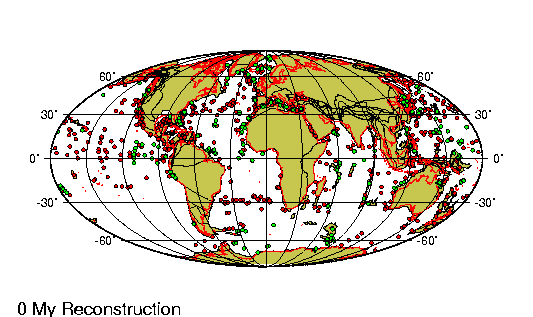
La historia evolutiva de los camélidos comienza en
América del Norte hace unos 45 millones de años.
Un pequeño antecesor de 30 centímetros de talla como
Protylopus petersoni, genera una serie de
especies con tendencia al aumento de tamaño hasta
que, hace alrededor de tres millones de años, un
gran camélido (Parazemelus) emigra a Asia por
el estrecho de Behring, diversificándose en las dos
especies de camellos que allí existen actualmente:
el camello con dos jorobas (Camelos bactrianus)
y el dromedario (Camelus dromedarius).
|
Aproximadamente para la misma época, Hemiauchenia,
otro camélido norteamericano parecido a la llama actual,
pasó por el istmo de Panamá (recién formado) hacia América
del Sur. La evolución en Sudamérica a partir de este
antecesor es poco conocida dado que el registro fósil es aun
incompleto.
Los
camélidos sudamericanos están emparentados con los camellos
que habitan en África y Asia. La diferencia morfológica
fundamental, es que éstos tienen joroba y son de mayor
tamaño. La diferenciación y especialización ha sido el
producto de un proceso evolutivo de millones de años.
Los
hallazgos de restos fósiles muestran que hace
aproximadamente 20 millones de años, los camélidos dominaban
las zonas planas de América del Norte. Un estudio detallado
de estos restos permitió clasificarlos en cuatro grupos con
características propias (Titanotylopus, Paracamelus,
Megatylopus y Hemiauchenia).
Debido al
enfriamiento de la tierra durante el Mioceno y el Plioceno,
las sabanas se incrementaron y los camélidos sufrieron
procesos selectivos que dieron lugar a adaptaciones a las
nuevas condiciones, como el alargamiento de sus patas, la
aparición de coronas en sus dientes, necesarias para
masticar los pastos.
A
continuación se citaran algunos géneros característicos que
dan luz sobre el origen y la biodiversidad de camelidos.
Protylopus.
Wortman, 1898.
Es un
género extinguido de camellos que vivió entre mediados y
finales del Eoceno, hace entre 45 y 40 millones de años en
América del Norte. Se trata del camello más antiguo que se
conoce y el más pequeño, con una longitud de 80 centímetros
y un peso de 26 kilogramos.
Por sus
dientes, es posible que se alimentase de hojas tiernas. Las
patas delanteras del Protylopus eran más cortas que
las traseras y contaban con cuatro dedos. Las patas traseras
también contaban con cuatro dedos, aunque la mayor parte del
peso se apoyaba en el tercero y el cuarto, por lo que se
podría elevar sobre las patas traseras para alimentarse,
como el generó actual.
Stenomylus.
Peterson, 1906
Es un
género extinto de pequeños camélidos. El Stenomylus
era mucho más pequeño que otros camélidos antiguos y
modernos, con una altura media de tan sólo 0,6 metros.
Al
contrario que los camélidos modernos, el Stenomylus
carecía de relleno en las pezuñas; se le ha comparado con el
berrendo de América del Norte y el género de África debido a
diferentes teorías sobre su biomecánica.
Aepycamelus.
MacDonald, 1956.
Es un
género extinto de mamífero artiodáctilo de la familia
Camelidae también conocida como Alticamelus en
literatura científica, que vivió en el Mioceno entre 20.6 a
4.9 millones de años.
Su nombre
se deriva del griego homérico αἰπύς, "alto y empinado" y κάμελος - "camello"; por lo que traduce "camello
alto"; su sinónimo Alticamelus tiene un significado
similar en latín. Aepycamelus caminaba sólo sobre sus
dedos. A diferencia de las primeras especies de camélidos,
estos poseían almohadillas acolchadas en las patas como las
de los camellos modernos.
 |
Aepycamelus
rondaba las praderas de Norteamérica (Colorado,
etc.). Era un animal bastante especializado. Su
cabeza era relativamente pequeña comparada con el
resto del cuerpo, con un cuello muy alargado, dando
como resultado un alargamiento de las vértebras
cervicales a la manera de las jirafas, con patas
largas y similares a zancos, con las articulaciones
del codo y la rodilla al mismo nivel. Su altura
máxima de la parte alta de la cabeza hasta el suelo
pudo haber sido de 3 metros |
Su extraña
estructura corporal ha dado una gran cantidad de información
acerca de su modo de vida y hábitos. Aepycamelus habitaba
planicies secas con grupos de árboles. Se presume que se debió
desplazar solo o en pequeños grupos, como las jirafas actuales,
y como estas, ramoneaba en los árboles altos. En este aspecto no
tenía competidores, sobreviviendo por un largo período de
tiempo, y perduró por la entera extensión de la época del
Mioceno, desapareciendo antes del inicio del Plioceno,
posiblemente debido a los cambios climáticos.
Gigantocamelus.
Barbour & Schultz, 1939.
|
 |
Es un
género extinto de mamíferos artiodáctilos de la familia
Camelidae que vivió durante el período comprendido entre
el Plioceno y el Pleistoceno tardío en Norteamérica,
desde hace 4.9 millones hasta 300,000 años, existiendo
por aproximadamente 4.68 millones de años.
Gigantocamelus
fue nombrada por Barbour y Schultz (1939), siendo su
especie tipo Gigantocamelus fricki. |
Posteriormente
fue considerada un sinónimo de Titanotylopus. Fue
asignada a la familia Camelidae. Se han encontrado fósiles de
Gigantocamelus en Texas, Nebraska, el oeste de Oregón y la
Sierra Nevada, en Nevada, California.
Titanotylopus
Barbour & Schultz, 1934.
Es un género
extinto de mamíferos artiodáctilos de la familia Camelidae que
vivió durante el período comprendido entre el Mioceno reciente y
el Pleistoceno tardío en Norteamérica, hace entre
4,9 millones de años
a 300,000 años
 |
Titanotylopus
se distingue por sus largos caninos superiores y la
reducción en los huesos lacrimales, además de las
relativamente amplias segundas falanges de sus patas, a
diferencia de sus contemporáneos como Camelops,
lo que sugiere que tenía patas con almohadillas como sus
parientes actuales.
La
especie Titanotylopus spatulus se caracterizaba
por sus amplios dientes incisivos espatulados. Se ha
hallado en Grand View (Idaho), Red Light, Condado de
Hudspeth, Donnelly Ranch, White Rock, Mullen II
(Kansas), Sandahl Local Fauna (Nebraska) y Vallecito
Creek, Colorado, Ranch, Arizona en Estados Unidos. |
Titanotylopus
poseía extremidades grandes y largas, un neurocráneo
comparativamente pequeño y una pendiente convexa entre los ojos.
Su altura promedio era de 3.5 metros. Al igual que el camello
moderno, probablemente poseía una joroba, evidencia de esto son
las largas apófisis espinosas de las vértebras torácicas.
Hemiauchenia. Gervais &
Ameghino, 1880.
Es una de las
especies del género extinto Hemiauchenia. Este camélido
de aspecto similar a una gran llama vivió en la porción austral
de América del Sur. Se extinguió con la llegada de los primeros
seres humanos.
 |
Por
otro lado, los Hemiauchenios se originaron de los
pliauchenia que aparecieron entre 9 y 11 millones
de años atrás en las praderas de América del norte.
Dando origen al género hemiauchenia hace 10
millones de años. Algunas especies de este género
migraron hacia el sur, hace aproximadamente 3 millones
de años, durante la transición del Plioceno al
Pleistoceno, (casi en la misma época en que los
antecesores de la tribu camelini emigraron a Asia
por el estrecho de Bering), pasando por el istmo
centroamericano, invadiendo las planicies y pampas de
Sudamérica. |
Tenía
costumbres corredoras, prefiriendo ambientes de pastizales
abiertos. Sus hábitos tróficos eran herbívoros, especialmente
ramoneadores. Los restos de esta especie fueron exhumados de
sedimentos cuaternarios cronoestimados entre los 781 mil y los
12 mil años aP, correspondientes al Pleistoceno Medio y Tardío,
hasta el límite con el Holoceno.
Hace unos diez
mil años, los camélidos se extinguieron en América del Norte.
Las causas de este fenómeno están aún en debate: algunos autores
opinan que se debería a cambios climáticos y otros al arribo del
predador más eficiente, el hombre. En América del Sur
desaparecen los Hemiauchenia y los Paleolama, y quedan solamente
los guanacos del género Lama y la vicuña del género
Vicugna. De las seis especies actuales, dos se encuentran en
Asia y cuatro en América del Sur.
Bibliografía sugerida.
Lamo, Daniel
(2011). Camélidos Sudamericanos: Historia, usos y sanidad
animal. Senasa.
Félix Pérez-Lorente; Cayetano Herrero; Emilio Herrero; Plinio
Montoya (2009). «Paracamelichnum Jumillensis n.ichnogen.
n.ichnosp., Upper Miocene Camelidae Ichnites from the Hoya de la
Sima site (Murcia, Spain)». Ichnos (Universidad de La Rioja) 16
(3): 13.
Peng Cui; Rimutu Ji; Feng Ding; Dan Qi; Hongwei Gao; He Meng;
Jun Yu; Songnian Hu et al. (18 de julio de 2007).
«A complete mitochondrial genome sequence of the wild two-humped
camel (Camelus bactrianus ferus): an evolutionary history of
camelidae».
BMC Genomics
8 (241).
Pinto Jiménez, Chris Evelyn; Martín Espada, Carmen; Cid
Vázquez, María Dolores (2010). «Camélidos sudamericanos:
clasificación, origen y características». Revista complutense de
ciencias veterinarias 4 (1): 23-26.
Wilson, Don; Reeder, DeeAnn, eds. (2005). «Lama glama». Mammal
Species of the World (en inglés)
(3ª edición). Baltimore: Johns Hopkins University Press, 2 vols.
(2142 pp.).
Los comienzos de la
Paleobotánica.
Por Mariano
Magnussen Saffer. Grupo Paleo. Museo Municipal Punta Hermengo de
Miramar.
marianomagnussen@yahoo.com.ar.
Tomado de;
Magnussen Saffer,
Mariano. 2016.
Los
comienzos de la Paleobotánica.
Paleo Revista Argentina de
Paleontología. Boletín Paleontológico. Año XIV. 147: 26-27.
La
paleobotánica es una disciplina compartida por la Botánica y la
Paleontología que estudia los restos de vegetales que vivieron
en el pasado. También contempla el uso de los restos para la
reconstrucción de ambientes antiguos y la historia de la vida.
Incluye el estudio de los fósiles de las plantas terrestres, y
los autótrofos marinos como las algas.
Los fósiles
vegetales son restos de individuos que se han conservado en las
rocas sedimentarias por un proceso fisicoquímico denominado
fosilización. Por lo general sólo se conservan las partes más
duras, siendo excepcional que se conserven también las partes
más blandas, gracias a esto es posible que lleguen hasta
nosotros algunas de las partes indispensables a la hora de
clasificar una planta, estas son: las hojas, parte de sus
tallos, semillas y resina fósil.
 |
Es
infrecuente hallar los restos completos de un individuo,
con todos sus elementos, lo corriente es encontrar en el
sitio una variedad de tipos de hojas, semillas, madera
en estado fósil lo que hace muy difícil el relacionar
unas con otras.
Esto ha
dado lugar a que cada elemento (raíz, tallo, hoja, etc.)
sea clasificado en forma independiente como si fueran
especies distintas. Las categorías —y por extensión los
componentes de las mismas— de esta sistemática paralela
basada en elementos aislados se denominan parataxones,
para diferenciarlas de los taxones formales. |
Esta disciplina
nos permite deducir el clima de entonces, su evolución y la
influencia sobre otros organismos.
Durante el
siglo XIX esta disciplina encuentra su origen. En 1828 el hijo
de Alexandre Brongniart, el botánico Adolphe Brongniart, publicó
la introducción a un trabajo más extenso sobre la historia de
las plantas fósiles. Adolphe Brongniart concluyó que la historia
de las plantas más o menos se podía dividir en cuatro partes.
El primer
período se caracterizaba por la presencia de criptógamas. El
segundo período se caracterizaba por la aparición de las
coníferas. El tercer período incluía el surgimiento de las
cícadas, y el cuarto el desarrollo de las plantas con floración
(tales como las dicotiledóneas).
Las
transiciones entre cada uno de estos períodos estaban marcadas
por discontinuidades bruscas en el registro fósil, con cambios
más graduales dentro de los períodos. El trabajo de Brongniart
es la base de la paleobotánica y reforzó la teoría de que la
vida en la tierra tenía una historia larga y compleja, y los
diferentes grupos de plantas y animales aparecían en orden
sucesivo.
|
 |
También
apoyó la idea de que el clima terrestre había cambiado
con el tiempo, dado que Brongniart concluyó que el
registro fósil de plantas demostraba que el clima del
norte de Europa debía haber sido tropical durante el
Carbonífero.
La
atención cada vez mayor que se prestaba a las plantas
fósiles durante las primeras décadas del siglo
xix
desencadenaría un cambio significativo en la
terminología para el estudio de la vida pasada.
|
El editor de la
influyente revista científica francesa Journal de Physique,
un estudiante de de Cuvier llamado Henri Marie Ducrotay de
Blainville, acuñó el término "paleozoología" en 1817 para
referirse a la obra que Cuvier y otros estaban haciendo para
reconstruir animales extintos a partir de huesos fósiles.
Sin embargo,
Blainville comenzó a buscar un término que pudiera referirse al
estudio tanto de fósiles de animales como de restos vegetales.
Después de probar algunas alternativas sin éxito, se le ocurrió
"palaeontología" en 1822. El término de Blainville para el
estudio de los organismos fosilizados adquirió popularidad
rápidamente y se derivó en el término actual "paleontología".
Bibliografia sugerida;
Bowler, Peter J. (2003).
Evolution:The History of an Idea. University of California
Press.
Cadbury, Deborah (2000), The Dinosaur Hunters: A True Story of
Scientific Rivalry and the Discovery of the Prehistoric World,
Fourth Estate.
Desmond, Adrian (1975). "The Discovery of Marine Transgressions
and the Explanation of Fossils in Antiquity". American Journal
of Science, Volume 275.
Larson, Edward J. (2004). Evolution: the remarkable history of
scientific theory. Modern Library.
Everhart, Michael J. (2005). Oceans of Kansas: A Natural History
of the Western Interior Sea. Indiana University Press.
Palmer, Douglas (2005) Earth Time: Exploring the Deep Past from
Victorian England to the Grand Canyon. Wiley, Chichester.
Wikipedia.
Durante las primeras décadas
del siglo XIX la paleontología comenzó su carrera.
Por Mariano
Magnussen Saffer. Grupo Paleo. Museo Municipal Punta
Hermengo de Magnussen Saffer,
Mariano. 2016.
Durante
las primeras décadas del siglo XIX la paleontología comenzó
su carrera.
Paleo Revista Argentina de
Paleontología. Boletín Paleontológico. Año XIV. 146: 24-26.Miramar.
Fundación Argentavis.
marianomagnussen@yahoo.com.ar. Tomado de;
Durante los
primeros 30 años del siglo XIX, la paleontología comenzó a
tomar un formato mas parecido al actual, con muestras
comparativas y con nuevos términos revolucionarios para la
época.
En 1808,
Cuvier identificó un fósil encontrado en Maastricht como un
reptil marino gigante que más tarde sería nombrado
Mosasaurus. También identificó a partir de un dibujo
otro fósil encontrado en Baviera como un reptil volador que
denominó llamó Pterodactylus.
Especuló,
con base en los estratos en los que se encontraron estos
fósiles, que los grandes reptiles habían vivido antes de lo
que él llamaba "la era de los mamíferos". Las
especulaciones de Cuvier serían confirmadas por una serie de
hallazgos que se hicieron en Gran Bretaña en el transcurso
de las siguientes dos décadas. Mary Anning, una
coleccionista de fósiles profesional desde los once años,
recolectó los fósiles de diversos reptiles marinos de los
estratos marinos del Jurásico en Lyme Regis.
 |
Entre estos fósiles se incluía el primer esqueleto
de ictiosaurio que fue reconocido como tal,
recolectado en 1811, y los dos primeros esqueletos
de plesiosaurio encontrados en 1821 y 1823. Muchos
de sus descubrimientos serían descritos
científicamente por los geólogos William Conybeare,
Henry De la Beche, y William Buckland.
Fue
Anning quien observó que los objetos pétreos
conocidos como "piedras bezoar" se encontraban a
menudo en la región abdominal de los esqueletos de
ictiosaurios, y señaló que si se rompían dichas
piedras a menudo contenían huesos de peces y escamas
fosilizados, incluso huesos de pequeños ictiosaurios.
|
Esto la llevó a
sugerirle a Buckland que estaban tratando con heces fosilizadas,
que él nombró coprolitos, y que utilizó para entender mejor las
cadenas tróficas antiguas.
En 1824,
Buckland descubrió y describió una mandíbula inferior de los
depósitos del Jurásico en Stonesfield. Determinó que el hueso
pertenecía a reptil carnívoro terrestre al que llamó
Megalosaurus. Ese mismo año Gideon Mantell se dio cuenta de
que algunos dientes grandes que había encontrado en 1822 en
rocas del Cretácico en Tilgate pertenecían a un reptil herbívoro
terrestre gigante. Lo nombró Iguanodon, debido a que sus
dientes se asemejaban a los de una iguana. Este hecho condujo a
Mantell a publicar en 1831 un influyente artículo titulado "La
era de los reptiles" en el que se resumía la evidencia de que
existió un largo período de tiempo durante el cual la Tierra se
estuvo llena de grandes reptiles.
 |
Dividió
esta época basándose en los estratos de suelo en el que
los distintos tipos de reptiles aparecieron por primera
vez, es decir, en tres intervalos que se anticiparon a
los períodos modernos del Triásico, Jurásico y
Cretácico. En 1832, Mantell descubriría en Tilgate un
esqueleto parcial de un reptil acorazado al que denominó
Hylaeosaurus. En 1841 el anatomista Inglés
Richard Owen crearía un nuevo orden de reptiles, al que
nobró Dinosauria, para Megalosaurus, Iguanodon
y Hylaeosaurus. |
La evidencia de
que grandes reptiles habían habitado la Tierra en el pasado
causó revuelo y entusiasmo en la comunidad científica, e incluso
entre algunos sectores del público general. Buckland describió
la mandíbula de un pequeño mamífero primitivo,
Phascolotherium, que se encontró en los mismos estratos que
el Megalosaurus. Este descubrimiento, conocido como el
mamífero de Stonesfield, fue una anomalía profundamente
discutida.
En un
principio, Cuvier pensó que era un marsupial, pero Buckland se
dio cuenta más tarde de que era un mamífero placentario
primitivo. Debido a su pequeño tamaño y la naturaleza primitiva
del espécimen, Buckland no consideraba que se invalidaba el
patrón general de la era de los reptiles, sino que indicaba los
animales más grandes y visibles habían sido reptiles en vez de
mamíferos.
Bibliografia sugerida;
Bowler, Peter J. (2003).
Evolution:The
History of an Idea. University of California Press
Cadbury, Deborah (2000), The Dinosaur Hunters: A True Story of
Scientific Rivalry and the Discovery of the Prehistoric World,
Fourth Estate.
Desmond, Adrian (1975). "The Discovery of Marine Transgressions
and the Explanation of Fossils in Antiquity". American Journal
of Science, Volume 275.
Larson, Edward J.
(2004). Evolution: the remarkable history of scientific theory.
Modern Library.
Everhart, Michael J.
(2005). Oceans of Kansas: A Natural History of the Western
Interior Sea. Indiana University Press.
Palmer, Douglas (2005) Earth Time: Exploring the Deep Past from
Victorian England to the Grand Canyon.
Wiley,
Chichester.
Wikipedia.
El límite K/T y la gran extinción masiva del
Cretácico-Terciario.
Por
Mariano Magnussen Saffer. Director de Grupo Paleo y Presidente
de la
Asociación de Amigos del Museo Municipal Punta Hermengo.
marianomagnussen@yahoo.com.ar.
Paleo,
Revista Argentina de Paleontología. Boletín Paleontológico.
Año 9. 64: 12-14.
El límite K/T
es una huella geológica, presente como una estrecha capa en los
estratos de la corteza terrestre, que data aproximadamente de
hace 65 millones de años. El nombre proviene de K,
abreviatura aplicada al periodo Cretácico (en alemán Kreide)
y T, abreviatura normalmente aplicada al periodo
Terciario (en alemán Tertiär). Se considera que el límite
K/T marca el final de la era Mesozoica y el comienzo de la era
Cenozoica. Según la hipótesis Alvarez, este límite se
corresponde con el impacto de un objeto extraterrestre que
provocó la Extinción masiva del Cretácico-Terciario
En 1980 un
grupo de investigadores liderados por el físico Luis Álvarez
(Premio Nobel) descubrieron, en las muestras tomadas por todo el
mundo de las capas intermedias entre los períodos Cretácico y
Terciario de hace 65 millones de años, una concentración de
iridio cientos de veces más alta que lo normal. El final del
cretáceo coincide con la extinción de los dinosaurios y de los
ammonites.
 |
Plantearon así la llamada "Hipótesis Álvarez" o
"Hipótesis de Álvarez", conforme la cual la extinción de
los dinosaurios y de muchas otras formas de vida habría
sido causada por el impacto de un gran meteorito contra
la superficie de la Tierra hace 65 millones de años. El
nombre de la hipótesis se debe a los dos científicos que
propusieron la hipótesis en 1980: Luis Álvarez y Walter
Álvarez (padre e hijo). |
Para demostrar
esta hipótesis, las investigaciones se centraron en encontrar
una capa en la corteza de la Tierra con niveles elevados de
iridio. Los niveles del iridio son generalmente más altos en
asteroides y otros objetos extraterrestres. La evidencia del
iridio fue descubierta anteriormente al descubrimiento del
cráter de Chicxulub
La extinción
masiva del Cretácico-Terciario fue un período de extinciones
masivas de especies hace aproximadamente 65 millones de años.
Corresponde al final del período Cretácico y el principio del
período Terciario. También se le conoce como extinción masiva
del límite K/T (del alemán Kreide/Tertiär Grenze), para
señalar la frontera entre el Cretácico-Terciario.
No se conoce la
duración exacta de este evento. Cerca del 50% de los géneros
biológicos desaparecieron, entre ellos la mayoría de los
dinosaurios. Se han propuesto muchas explicaciones a este
fenómeno; la más aceptada es que fue el resultado del impacto de
un Asteroide sobre la Tierra proveniente del espacio.
Durante la
década que siguió a la publicación del estudio, la hipótesis de
la extinción por el choque de un asteroide continuó siendo tema
de debate entre geólogos y paleontólogos.
Uno de las
mayores objeciones a esta hipótesis era que no se conocía un
cráter cuyas dimensiones correspondieran al tamaño calculado,
que debería tener entre 150 y 200 km de diámetro. Si bien no
sería imposible que la Tierra hubiera cambiado desde entonces
escondiendo una tal deformación, en 1990 se ubicaron indicios en
Haití de un tsunami de grandes proporciones que arrastró
residuos de iridio. Buscando estudios geológicos realizados
desde los años 1960 en adelante se pudo ubicar un cráter en
Chicxulub, en la península de Yucatán, con un diámetro de unos
180 km.
Para algunos
científicos, un problema de esta teoría es que la lectura de los
registros fósiles sugiere que la extinción masiva de hace 65
millones de años duró cerca de diez millones de años, lo que no
cuadra bien con que su causa fuera el impacto. Otros autores
sostienen que la extinción fue muy rápida para la mayor parte de
las especies. Es evidente que gran parte de estas discusiones
está condicionada por la escasez de restos fósiles en grupos
como los dinosaurios, si se calcula que la población al momento
de su extinción pudo ser de 10.000 millones de estos animales en
todo el mundo. Hasta el momento, la única zona conocida rica en
restos de dinosaurios con continuidad sedimentaria a través del
límite K/T es la Formación Hell Creek de Norteamérica, donde los
especialistas en dinosaurios llevan décadas discutiendo sobre si
su extinción fue catastrófica o se produjo gradualmente a lo
largo de los últimos 10 millones de años del Cretácico.
 |
A pesar
de la dificultad de encontrar series ricas en fósiles de
dinosaurios donde se haya registrado el límite K/T, una
buena aproximación al debate puede realizarse calculando
cómo varía su diversidad con el tiempo. En este sentido,
el equipo de D. E. Fastowsky publicó en 2004, en la
prestigiosa revista Geology, un trabajo donde
analizaban estadísticamente la base de datos más
completa que existe sobre la diversidad de restos óseos,
huevos, coprolitos y huellas de dinosaurios encontrados
en los cinco continentes. |
Dichos autores
concluyeron que, lejos de descender, la diversidad de géneros
fósiles relacionados con los dinosaurios dentro de los últimos
18,5 millones de años del Cretácico alcanzó un máximo justamente
durante los dos millones de años previos al límite K/T,
contradiciendo el aparente declive gradual que algunos autores
han defendido.
Los
foraminíferos planctónicos (organismos unicelulares marinos) son
otro grupo muy estudiado en relación con las extinciones del
límite K/T. Según los recientes resultados científicos de un
equipo internacional de investigadores liderados por Gerta
Keller (Universidad de Princeton, EE. UU.) y Thierry Adatte
(Universidad de Neuchâtel, Suiza), el cráter es 300.000 años más
antiguo que la lámina K/T (Cretácico-Terciario). Por el
contrario, otros estudios con foraminíferos planctónicos
llevados a cabo por el equipo de Jan Smit (Universidad Libre de
Amsterdam) o por equipo de micropaleontología de la Universidad
de Zaragoza (España), sostienen que el impacto meteorítico tuvo
lugar coincidiendo con el límite K/T.
El Paleoceno es
inmediatamente posterior a la extinción masiva del final del
Cretácico, conocido como límite KT (Cretácico-Terciario),
que marca la desaparición de los dinosaurios (la extinción
masiva del Cretácico-Terciario). La desaparición de los
dinosaurios dejó sin cubrir la mayoría de los nichos ecológicos
en todo el mundo, y el nombre de "Paleoceno" proviene del griego
(palaios), que significa "viejo", y (kainos), "nuevo" haciendo
referencia a la fauna que apareció durante la época, antes de
que los modernos órdenes de mamíferos surgieran en el Eoceno.
Bibliografía
Sugerida.
Alvarez, L. W. (1983).
«Experimental evidence that an asteroid impact led to the
extinction of many species 65 million years ago. Proceedings of
the National Academy of Sciences 80 (2): pp. pp. 627-642.
Hooker, J.J., "Tertiary to Present: Paleocene", pp. 459-465,
Vol. 5. of Selley, Richard C., L. Robin McCocks, and Ian R.
Plimer, Encyclopedia of Geology, Oxford: Elsevier Limited, 2005.
ISBN
0-12-636380-3
Magnussen Saffer,
Mariano (2011). Algunos vertebrados hallados cronológicamente en
el Cretácico de la Republica Argentina. Paleo, Revista Argentina
de Paleontología. Boletín Paleontológico. Año 9. 61: 16-27.
Molina, E.; Alegret, L., Arenillas, I.; Arz, J. A.,
Gallala, N., Grajales-Nishimura, J. M., Murillo-Muñetón, G. y
Zaghbib-Turld, D. (2009).
«The Global Boundary Stratotype Section and Point for the base
of the Danian Stage (Paleocene, Paleogene, "Tertiary",
Cenozoic): auxiliary sections and correlation» (en inglés).
Episodes 32 (2): 84-95.
Stephen Jay Gould, ed., The Book of Life (New York: W.W. Norton
& Company, 1993), p. 182.
Wikipedia.
|
Charles Robert Darwin. |
|
El Británico que puso la evolución en boca de todos. |
Fragmento del articulo publicado
originalmente: Magnussen Saffer, Mariano. (2008).
Charles
Robert Darwin. El Británico que puso la evolución en boca de
todos.
Paleo. Año 6 numero 31: 29 - 31.
marianomagnussen@yahoo.com.ar
Científico
británico que sentó las bases de la moderna teoría evolutiva, al
plantear el concepto de que todas las formas de vida se han
desarrollado a través de un lento proceso de selección natural.
Su trabajo tuvo una influencia decisiva sobre las diferentes
disciplinas científicas, y sobre el pensamiento moderno en
general.
Nació en Shrewsbury (Shropshire),
el 12 de febrero de 1809. Fue el quinto hijo de una acomodada y
sofisticada familia inglesa. Su abuelo materno fue el próspero
empresario de porcelanas Josiah Wedgwood; su abuelo paterno fue
el famoso médico del siglo XVIII Erasmus Darwin. Tras terminar
sus estudios en la Shrewsbury School en 1825, estudió Medicina
en la Universidad de Edimburgo. En 1827 abandonó la carrera e
ingresó en la Universidad de Cambridge con el fin de convertirse
en ministro de la Iglesia de Inglaterra. Allí conoció a dos
influyentes personalidades: el geólogo Adam Sedgwick y el
naturalista John Stevens Henslow. Este último no sólo ayudó a
Darwin a ganar confianza en sí mismo, sino que también inculcó a
su alumno la necesidad de ser meticuloso y esmerado en la
observación de los fenómenos naturales y la recolección de
especímenes. Tras graduarse en Cambridge en 1831, el joven
Darwin se enroló a los 22 años en el barco de reconocimiento HMS
Beagle como naturalista sin paga, gracias en gran
medida a la recomendación de Henslow, para emprender una
expedición científica alrededor del mundo.
Su trabajo como naturalista a bordo del Beagle le dió la
oportunidad de observar variadas formaciones geológicas en
distintos continentes e islas a lo largo del viaje, así como una
amplia variedad de fósiles y organismos vivos. En sus
observaciones geológicas, Darwin se mostró muy sorprendido por
el efecto de las fuerzas naturales en la configuración de la
superficie terrestre.
 |
En aquella época, la mayoría de los geólogos defendían la teoría
catastrofista, que mantenía que la Tierra era el resultado de
una sucesión de creaciones de la vida animal y vegetal, y que
cada una de ellas había sido destruida por una catástrofe
repentina, por ejemplo una convulsión de la corteza terrestre (véase
Geología: Siglos XVIII y XIX). Según esta teoría, el
cataclismo más reciente, el diluvio universal, había acabado con
todas las formas de vida no incluidas en el arca de Noé. Las
demás sólo existían en forma de fósiles. En opinión de los
catastrofistas, cada especie había sido creada individualmente y
era inmutable, es decir, no sufría ningún cambio con el paso del
tiempo.
Este punto de vista (aunque no la inmutabilidad de las especies)
había sido cuestionado por el geólogo inglés sir Charles Lyell
en su obra en dos volúmenes Principios de Geología
(1830-1833). |
Lyell sostenía que la superficie terrestre está
sometida a un cambio constante como resultado de fuerzas
naturales que actúan de modo uniforme durante largos periodos de
tiempo.
A bordo del Beagle, Darwin
descubrió que muchas de sus observaciones encajaban en la teoría
uniformitaria de Lyell. No obstante, durante su viaje por
Sudamérica, también observó gran diversidad de plantas, animales
y fósiles, y recogió gran número de muestras que estudió a su
regreso a Inglaterra. En las islas Galápagos, situadas frente a
la costa de Ecuador, observó especies estrechamente emparentadas
pero que diferían en su estructura y en sus hábitos
alimenticios, y concluyó que estas especies no habían aparecido
en ese lugar sino que habían migrado a las Galápagos procedentes
del continente. Darwin no se dio cuenta en ese momento que los
pinzones de las diferentes islas del archipiélago pertenecían a
especies distintas. Más tarde, ya en Inglaterra, llegaría a la
conclusión de que, cuando los pinzones llegaron al archipiélago
desde el continente encontraron gran variedad de alimento, y al
no tener competidores y estar aislados geográficamente,
sufrieron una rápida adaptación a los distintos ambientes; con
lo cual aparecieron nuevas especies que descendían todas ellas
de un antepasado común.
Tras su regreso a Inglaterra en 1836, Darwin comenzó a recopilar
sus ideas acerca del cambio de las especies en sus Cuadernos
sobre la transmutación de las especies. La explicación de la
evolución de los organismos le surgió tras la lectura del libro
Ensayo sobre el principio de la población (1798) del
economista británico Thomas Robert Malthus, que explicaba cómo
se mantenía el equilibrio en las poblaciones humanas. Malthus
sostenía que ningún aumento en la disponibilidad de alimentos
básicos para la supervivencia del ser humano podría compensar el
ritmo de crecimiento de la población. Este, por consiguiente,
sólo podía verse frenado por limitaciones naturales, como las
hambrunas o las enfermedades, o por acciones humanas como la
guerra.
Darwin aplicó de inmediato el razonamiento de Malthus a los
animales y a las plantas, y en 1838, había elaborado ya un
bosquejo de la teoría de la evolución a través de la selección
natural. Durante los siguientes veinte años trabajó sobre esta
teoría y otros proyectos de historia natural. Darwin disfrutaba
de independencia económica y nunca tuvo necesidad de ganarse la
vida. En 1839 se casó con su prima, Emma Wedgwood, y poco
después se instalaron en la pequeña propiedad de Down House, en
Kent. Allí tuvieron diez hijos, tres de los cuales murieron
durante la infancia.
Darwin hizo pública su teoría por primera vez en 1858, al mismo
tiempo que lo hacía Alfred Russel Wallace, un joven naturalista
que había desarrollado independientemente la teoría de la
selección natural. La teoría completa de Darwin fue publicada en
1859 como El origen de las especies por medio de la selección
natural. Este libro, del que se ha dicho que “conmocionó al
mundo”, se agotó el primer día de su publicación y se tuvieron
que hacer seis ediciones sucesivas.
 |
En esencia, la teoría de la evolución por selección natural
sostiene que, a causa del problema de la disponibilidad de
alimentos descrito por Malthus, los jóvenes miembros de las
distintas especies compiten intensamente por su
supervivencia. Los que sobreviven, que darán lugar a la
siguiente generación, tienden a incorporar variaciones
naturales favorables (por leve que pueda ser la ventaja
que éstas otorguen), al proceso de selección natural, y
estas variaciones se transmitirán a través de la
herencia. En consecuencia, cada generación mejorará en
términos adaptativos con respecto a las anteriores, y este
proceso gradual y continuo es la causa de la evolución de las
especies. La selección natural es sólo parte del amplio esquema
conceptual de Darwin. Introdujo también el concepto de que todos
los organismos emparentados descienden de antecesores comunes.
Además ofreció un respaldo adicional al antiguo concepto de que
la propia Tierra no es estática sino que está evolucionando. |
Las reacciones ante El Origen de las especies fueron
inmediatas. Algunos biólogos adujeron que Darwin no podía probar
su hipótesis. Otros criticaron su concepto de variación,
sosteniendo que ni podía explicar el origen de las variaciones
ni cómo se transmitían a las sucesivas generaciones. Esta
objeción en concreto no encontró respuesta hasta el nacimiento
de la genética moderna a comienzos del siglo XX (véase
Leyes de Mendel). Fueron muchos los científicos que siguieron
expresando sus dudas durante los ochenta años siguientes. Sin
embargo, los ataques a las ideas de Darwin que encontraron mayor
eco no provenían de sus contrincantes científicos, sino de sus
oponentes religiosos. La idea de que los seres vivos habían
evolucionado por procesos naturales negaba la creación divina
del hombre y parecía colocarlo al mismo nivel que los animales.
Ambas ideas representaban una grave amenaza para la teología
ortodoxa.
Darwin pasó el resto de su vida ampliando diferentes aspectos de
los problemas planteados en El Origen de las especies.
Sus últimos libros, entre los que se encuentran La variación
de los animales y de las plantas bajo la acción de la
domesticación (1868), La descendencia humana y la
selección sexual (1871), y La expresión de las emociones
en el hombre y en los animales (1872) eran exposiciones
detalladas sobre temas que sólo disfrutaban de un espacio
limitado en El origen de las especies. La importancia de
su trabajo fue ampliamente reconocida por sus coetáneos. Darwin
fue elegido miembro de la Royal Society (1839) y de la Academia
Francesa de las Ciencias (1878). Tras su muerte en Down, el 19
de abril de 1882 se le rindió el honor de ser enterrado en la
abadía de Westminster.
Bibliografía.
|
Castrodeza, Carlos. Ortodoxia
darwiniana y progreso biológico. Madrid: Alianza
Editorial, 1988. Obra de carácter divulgativo. |
|
Darwin, Charles R. Viaje de un
naturalista alrededor del mundo. 2 tomos, Madrid:
Ediciones Akal, 1983. Viaje que realizó el autor
alrededor del mundo. |
|
Darwin, Charles R. Autobiografía.
2 vols., Madrid: Alianza Editorial, 1984. Obra
autobiográfica de Darwin. |
|
Darwin, Charles R. El viaje del Beagle.
Barcelona: Editorial Labor, 1984. Viaje que realizó el
autor alrededor del mundo. |
|
Darwin, Charles. Origen de las
especies. 2ª ed., 1994. Madrid: Ediciones Akal.
Libro donde el autor fundamenta su teoría de la
evolución. |
|
Ghiselin, Michael T. El triunfo de
Darwin. Madrid: Ediciones Cátedra, 1983. Obra
divulgativa sobre la teoría de la evolución de Darwin. |
|
Howard, Jonathan. Darwin. Madrid:
Alianza Editorial, 1987. Obra biográfica sobre la figura
de Charles Darwin. |
|
Mayr, Ernest. Una larga controversia:
Darwin y el darwinismo. Barcelona: Editorial
Crítica, 1992. Obra analítica sobre Darwin y sus ideas. |
|
Microsoft Corporation, 2007. Charles
Robert Darwin."
Microsoft® Student 2008 [DVD].
|
|
Papp, Desiderio. Darwin. La aventura
de un espíritu. Madrid: Editorial Espasa-Calpe,
1983. Obra biográfica sobre la figura de Darwin. |
|
Ruse, Michael. La revolución
darwinista. Madrid: Alianza Editorial, 1983. Obra
divulgativa sobre la teoría de la evolución de Darwin. |
|
Darwin Charles Robert
Darwin Charles Robert Darwin Charles Robert Darwin
Charles Robert Darwin Charles Robert Darwin Charles |
|
|
200 aniversario de Charles
Darwin. Sus teorías nos insertaron en la naturaleza como una
especie más.
Publicado
por José Luís Santos Fernández. Tomado
de
Terrae Antiqvae.
Una herencia científica en
plena evolución. En el bicentenario de Darwin, cinco científicos
de distintas ramas de la biología valoran la pervivencia del
modelo evolutivo formulado por el inglés.
El día 12 de Febrero de 2009 se cumplieron 200 años del
nacimiento del naturalista inglés Charles Darwin. En noviembre
se conmemorará también el 150 aniversario de la publicación de
su obra El origen de las especies, la primera que expuso de
manera específica, exhaustiva y fundamentada la noción, ya
intuida por otros científicos en el siglo XIX, de que las
especies evolucionan a partir de ancestros comunes en un proceso
continuo y gradual que las permite adaptarse a los embates de su
medio.
La originalidad del británico
consistió en proponer mecanismos directores de la evolución,
sobre todo la llamada selección natural. Antes de Darwin, la
aproximación más cercana a un esquema evolutivo era la formulada
por el francés Jean-Baptiste Lamarck, para quien eran los
organismos individuales, no las especies, los que se adaptaban a
la fuerza a los cambios en el medio y legaban esas variaciones a
sus descendientes. En un ejemplo clásico del lamarckismo, la
jirafa habría surgido por la necesidad de estirar el cuello para
alcanzar las hojas en las copas altas de los árboles.
Tanto Darwin como Lamarck
desconocían la genética, el ADN y los mecanismos de la herencia,
por lo que la hipótesis del francés no resultaba tan
descabellada como hoy. Pero al contrario que el lamarckismo, el
modelo de Darwin era fácilmente compatible con lo que a diario
observaban los criadores de animales domésticos en sus procesos
de selección de razas, aunque la herencia continuase siendo una
caja negra para la ciencia de la época.
En el contexto de entonces,
donde los descubrimientos científicos despuntaban en el magma de
la crisis de fe de la sociedad victoriana, había ya una cierta
apertura hacia las interpretaciones de la historia natural que
se apartaban de las escrituras sagradas. Pese a ello, Darwin
esperó casi un cuarto de siglo después de su viaje de
exploración y recogida de datos en el navío HMS Beagle hasta
publicar finalmente su modelo, y lo hizo en parte presionado por
el descubrimiento de que el galés Alfred Russell Wallace había
llegado a similares conclusiones de manera independiente.
Pero aún persistía un tabú: la
posición del ser humano como algo esencialmente diferente y
superior al resto de la naturaleza. Aunque la obra pionera de
Darwin no indagaba en el ser humano, la aplicación del modelo
era inmediata y evidente. Antes de que el naturalista abordase
el asunto años más tarde en El origen del hombre, los círculos
científicos ya discutían una paternidad común para los humanos y
los simios, lo que soliviantó a la religiosidad del momento y
multiplicó las caricaturas que encastraban la cabeza de Darwin
en un cuerpo simiesco.
Dos siglos después, las
hipótesis de Darwin gozan de buena salud. Su propuesta básica se
ha contrastado en la naturaleza, se ha experimentado en el
laboratorio y se ha simulado con modelos informáticos.
Entretanto, la figura y su obra han sufrido innumerables asedios
y manipulaciones. Voces acientíficas propagan presuntas dudas
sobre su validez, otras falsean consanguineidades con el
nazismo, y el ateísmo militante lo enarbola como bandera.
Mientras el darwinismo se debate en contextos sociales y
religiosos que su autor nunca exploró, expertos como el
hispano-estadounidense Francisco J. Ayala (una de las máximas
autoridades mundiales en evolución) se empeñan inútilmente en
reclamar que se deje a la ciencia lo que es de la ciencia.
Darwin, desde luego, no era
infalible. Aunque hoy ningún biólogo reconocido duda de que las
especies evolucionen y que al menos uno de sus motores es la
selección natural, la biología evolutiva maneja modelos que han
adelantado en varias generaciones al darwinismo original.
Incluso la representación del viaje de las especies en el tiempo
como un árbol, algo que en su día fue revolucionario y que hoy
parece incuestionable, es cuestionado en favor de un esquema más
transversal en forma de red. Pero de algo no hay duda: la
semilla de Darwin fructificó en un árbol del que brotaron muchas
de las ramas de la biología moderna.
Cuatro
preguntas para cinco científicos
"Fundó la ciencia del hombre" Juan Luis Arsuaga.
Paleoantropología
Codirector de Atapuerca, Centro UCM-ISCIII de Evolución Humana
1 ¿Cómo pervive la herencia de Darwin en su disciplina?
Se puede decir que él la fundó, cambió las reglas del juego.
Antes de él no existía una ciencia del hombre. Sus teorías nos
insertaron en la naturaleza como una especie más.
2 ¿En qué aspectos su modelo era incompleto o erróneo?
No hay que momificar a Darwin. Él abrió las puertas, pero no
cerró ninguna. Hizo las preguntas que los demás seguimos
respondiendo. Por ejemplo, propuso dos mecanismos evolutivos, la
selección natural y la sexual. Hoy continuamos discutiendo qué
papel ha jugado cada uno.
3 ¿Qué destacaría de su huella social?
La evolución es indiscutida dentro del mundo científico, las
alternativas son ideológicas. El creacionismo ya no cabe ni
siquiera en el humanismo ni en la cultura; es el no pensamiento.
Es cuestión de tiempo que desaparezca, como ocurrió con
Copérnico. Hoy nadie cree en el geocentrismo.
4 ¿Qué le preguntaría hoy a Darwin si tuviera ocasión?
En qué momento vislumbró la evolución, si fue, como creemos, a
su vuelta del Beagle, entre marzo y junio de 1837.
"El peso del dogma es excesivo" Carlos Duarte. Ecología
Instituto Mediterráneo de Estudios Avanzados CSIC-UIB
1.
Ha tenido un peso crucial en el desarrollo de la ecología de
poblaciones. Por su condición de dogma, a veces el peso de esta
herencia es excesivo y detrae de una exploración más libre de
los resultados.
2.
Ignoraba la posibilidad de saltos evolutivos bruscos por dos
procesos: los cooperativos en lugar de tanto énfasis en los
competitivos, y la transferencia genética transversal entre
organismos situados en ramas alejadas del árbol de la vida.
3.
El darwinismo social es uno de los fundamentos del capitalismo
más salvaje, renacido en la fórmula del neoconservadurismo cuyas
consecuencias padecemos en este momento.
4.
Estoy a punto de embarcarme en el Hespérides rumbo a la
Antártida. Le preguntaría por su relación con el capitán Fitz-Roy.
La colaboración entre armada y científicos es fuente de éxitos,
pero hay algún desencuentro. Su experiencia podría enseñarnos
algo.
"Los genes aportaron otra prueba" Antonio Barbadilla. Genética
Universidad Autónoma de Barcelona
1.
Es imprescindible. Consiguió que ningún científico dudase de la
evolución. No podríamos entender la genética y la genómica sin
él. Quizá lo más interesante es que lo que él vio a nivel
morfológico se confirma en los genes, aportando otra prueba
independiente.
2.
Al no conocer la genética, no entendía cómo los caracteres no
se diluían al mezclarse en las sucesivas generaciones. Esto le
producía cierta inseguridad. Pero no hemos hecho más que ampliar
sus ideas mediante la genética. Es sorprendente cómo la
filogenia del ADN nos revela las capas de la evolución a través
de los genes que compartimos.
3.
Es alarmante el desfase entre científicos y ciertos sectores de
la sociedad anclados en el creacionismo. Es la gran paradoja de
la cultura moderna.
4.
Si habría imaginado que la genética vindicaría su teoría. Le
habría entusiasmado, porque no sabía nada de este campo.
"Le habría planteado la panspermia" Ricardo Amils.
Astrobiología
Universidad Autónoma de Madrid y Centro de Astrobiología
CSIC-INTA
1.
Su primera piedra en los conceptos evolutivos ha madurado en la
búsqueda de relojes moleculares que han facilitado la ordenación
de los microorganismos.
2.
Hoy sabemos que los mecanismos evolutivos son mucho más
complejos, sobre todo en bacterias, pero eso no le resta
importancia. No hemos avanzado mucho más sobre el papel de las
condiciones ambientales en la selección.
3.
En la sociedad culta, Darwin es incuestionable. Pero fuera de
esta torre de marfil, mucha gente cree en el diseño inteligente
o el creacionismo porque no requiere dar muchas explicaciones.
Deberíamos salir más a menudo para divulgar, es nuestra
responsabilidad social como científicos.
4.
Dónde colocaría los virus en su modelo evolutivo, o qué le
parece la panspermia [origen extraterrestre de la vida] para
explicar la rápida instauración de la vida en nuestro planeta.
"Le debemos los modelos animales" Juli Peretó. Bioquímica
Instituto Cavanilles de Biodiversidad y Biología Evolutiva
1.
Hay una unidad de procesos metabólicos y bioquímicos en los
seres vivos que nos permite emplear animales como modelos. Si no
fuera por Darwin, no tendría sentido utilizar un ratón como
modelo de enfermedad humana.
2.
La bioquímica no existía en su tiempo. Y a pesar de eso, Darwin
está en los cimientos de la bioquímica moderna, que ha ampliado
sus ideas al mundo subcelular.
3.
Lo curioso es que los creacionistas lo son sólo el fin de
semana. De lunes a viernes se ponen en manos de Darwin: se
tratan infecciones o aplican remedios contra plagas agrícolas,
cosas que existen gracias a nuestro conocimiento de la
evolución.
4.
Por qué, al sugerir la evolución previa a animales y plantas,
nunca pensó en los microorganismos. Y también, qué sintió al
recibir la carta de Wallace en la que descubría lo mismo que él.
Completando a Charles Darwin
La tectónica, la oceanografía o el clima están dando respuesta a
los interrogantes pendientes sobre la evolución - Los nuevos
hallazgos cierran lagunas en el 200º aniversario del científico.
Una crítica clásica contra
Darwin es que, pese a haber titulado su libro El origen de las
especies (1859), justo no aclaró cómo se originaban las
especies. La selección natural -el mecanismo evolutivo
descubierto por el naturalista- se basa en la acumulación
gradual de pequeños cambios, mientras que las especies suelen
ser entidades discretas y bien definidas: vemos leones y tigres,
no una escala Pantone de leotigres. La investigación reciente,
sin embargo, ha aclarado muchos puntos del problema de la
especiación, o generación de nuevas especies, y ha confirmado
que la especiación tiene una relación directa con la selección
natural darwiniana. También han revelado unos principios
generales que hubieran resultado sorprendentes para el padre de
la biología moderna.
"La competencia por los recursos, las carreras de armamentos
entre predadores y presas y otros factores biológicos dan forma
a los ecosistemas locales durante periodos cortos", dice el
evolucionista Michael Benton, de la Universidad de Bristol.
"Pero son factores externos como el clima, la oceanografía y la
tectónica continental los que explican las pautas de la
evolución a gran escala". Benton es el autor de uno de los cinco
artículos con que la revista Science celebra hoy el 200º
aniversario del nacimiento de Charles Darwin (12 de febrero de
1809-19 de abril de 1882).
La idea de que la competencia entre seres vivos es el principal
motor de la evolución arranca del propio Darwin y suele ser la
preferida por los biólogos. Se la conoce como la hipótesis de la
reina roja, por el personaje de Lewis Carroll que le dice a
Alicia en A través del espejo: "En este país tienes que correr
todo lo que puedas para permanecer en el mismo sitio".
El paradigma de la reina roja son las carreras de armamentos
entre predador y presa: los conejos corren cada vez más para
escapar de los zorros, lo que fuerza a los zorros a correr cada
vez más para seguir comiendo lo mismo que antes; las corazas de
las presas se hacen cada vez más duras y las pinzas de sus
predadores cada vez más fuertes, con lo que todos corren lo más
que pueden para que todo permanezca en el mismo sitio.
El problema es que la evolución a gran escala no permanece en el
mismo sitio como Alicia. Los modelos del tipo reina roja, según
Benton, no explican que los seres vivos se hayan hecho más
complejos en la historia del planeta, ni que hayan colonizado
nuevos espacios (como la tierra firme), ni que ciertos linajes
concretos hayan brotado en explosiones evolutivas de radiación
de nuevas especies. "Todas estas cosas han ocurrido muchas veces
en los últimos 500 millones de años", afirma el científico
británico.
La razón hay que buscarla en la geología, y algunos ejemplos son
bien conocidos. Desde que el supercontinente Pangea empezó a
quebrarse hace 250 millones de años, el baile de sus fragmentos
por la corteza terrestre ha tenido un efecto decisivo. La
biología alienígena de Australia -ornitorrincos, canguros,
koalas, wombats, emús, cucaburras- y de Suramérica -llamas,
anacondas, pirañas, vicuñas, tapires- se debe a que ambos
territorios han sido islas durante casi 100 millones de años.
El sentido común no es la mejor guía para averiguar las
relaciones de parentesco entre las distintas especies. El damán,
un animalillo africano al que cuesta distinguir de una rata, se
agrupa con el elefante en una gran rama evolutiva de los
mamíferos, la de los afroterios. Las personas, los delfines y
las vacas nos apiñamos junto a las ratas propiamente dichas en
la segunda rama (los boreoterios), dejando la tercera (los
desdentados) para el armadillo y el oso hormiguero.
La razón es que los mamíferos originales se dividieron
físicamente en tres grupos hace 100 millones de años, cuando las
actuales África, Eurasia y Suramérica se escindieron de un
continente único.
En los últimos años, los geólogos también han encontrado fuertes
correlaciones entre la diversidad del plancton -los organismos
microscópicos que flotan en el mar- y la temperatura del agua en
esa época. El enfriamiento oceánico de los últimos 70 millones
de años, por ejemplo, se asocia a una gran radiación de especies
de foraminíferos, los principales microfósiles marinos. En
general, las fases de calentamiento por las que ha pasado el
planeta se han caracterizado por una menor riqueza de géneros, y
de familias enteras, de seres vivos.
Si la competencia entre seres vivos es la reina roja, la
evolución guiada por las condiciones externas se conoce como la
hipótesis del "bufón de corte". Los bufones sólo pretendían
complacer a los poderosos, y jamás cambiaban sus números a menos
que se vieran forzados por una catástrofe (como una guerra o un
cambio de régimen). Si la reina roja es la idea preferida por
los biólogos, el bufón de corte es la favorita de los geólogos,
como parece lógico. Y es el motor del cambio que parece
predominar a las escalas evolutivas, de 100.000 años para arriba
en el tiempo, y de especie para arriba en la taxonomía, la
ciencia que clasifica a los seres vivos en una jerarquía de
especies, géneros, familias, órdenes, clases, filos y reinos.
La cuestión de la reina roja tiene mucha relevancia para el
problema estrella de la biología evolutiva: la explosión
cámbrica, la gran dificultad que atormentó a Darwin hace un
siglo y medio. La Tierra tiene 4.500 millones de años, y los
primeros microbios aparecieron poco después (hay evidencias
fósiles de 3.500 millones de años). Pese a ello, la explosión de
la vida animal sólo ocurrió al empezar el periodo Cámbrico, hace
543 millones de años. La evolución tardó poco en inventar a los
animales, aunque tardó 3.000 millones de años en ponerse a ello.
Ésta es la versión moderna del dilema de Darwin.
"Creo que la explosión cámbrica es un excelente ejemplo de
evolución por el modelo del bufón de corte", confirma Benton a
EL PAÍS. "Es un caso en que el cambio dramático del entorno
físico tiene un profundo efecto en la evolución. Esto no tiene
nada que ver con sugerir que la selección natural es errónea, o
que Darwin se equivocó. Se trata simplemente de que los cambios
dramáticos e inesperados, como el que ocurrió entonces, pueden
abrumar a los procesos normales de la selección natural y poner
a cero el reloj evolutivo, como solía decir Steve Gould".
Stephen Jay Gould fue un destacado (y polémico) evolucionista
norteamericano hasta su muerte en 2002.
El periodo anterior al Cámbrico (de 1.000 a 543 millones de años
atrás) se llama Neoproterozoico, de mote "precámbrico", e
incluye las más brutales glaciaciones conocidas por los
geólogos, como la Sturtian y la Marinoan. Algunos científicos
creen que fue una era de bola de nieve planetaria (snowball
earth), en la que los casquetes polares cubrían incluso el
ecuador terrestre.
Antes de esa era del hielo, los niveles de oxígeno en la
atmósfera eran muy bajos, inferiores al 1% de la concentración
actual, como habían sido en los 3.000 millones de años
anteriores. La última de las grandes glaciaciones precámbricas,
la Marinoan, terminó hace 635 millones de años, y los últimos
datos indican que los primeros animales, las esponjas, ya habían
evolucionado para entonces. Y los datos indican que el fondo
marino no estuvo bien oxigenado hasta los tiempos de la
explosión cámbrica. Si la biología tardó 3.000 millones de años
en inventar a los animales, la razón parece ser que la geología
no se lo permitió antes.
La mosca Drosophila ha resultado un modelo muy útil para
estudiar los fundamentos genéticos de la especiación. Por
ejemplo, la especie americana Drosophila pseudoobscura se separó
hace 200.000 años en dos subespecies llamadas USA y Bogotá. Como
los caballos y los burros, las moscas USA y Bogotá pueden
cruzarse, pero sus hijos son estériles. En casos de especies más
divergentes, los hijos suelen ser no ya estériles, sino
directamente inviables. El punto es que la genética de la mosca
permite hallar los genes exactos que son responsables de la
esterilidad o de la inviabilidad.
Los resultados apuntan a muy pocos genes, y varios están
relacionados con el transporte nuclear, el intercambio de
materiales entre el núcleo y el resto de la célula. Dos de los
genes de la especiación son Nup96 y Nup160, componentes del poro
nuclear que comunica al núcleo con su entorno, y otro es RanGAP,
que regula el mismo proceso. No hay ninguna razón a priori para
que la especiación esté relacionada con un mecanismo tan
concreto como el transporte nuclear, y estos resultados son
inesperados en ese sentido.
Pero estos genes también tienen relación con un fenómeno que
lleva décadas siendo un sospechoso central para los genetistas
interesados en la especiación. Se llama impulso meiótico (meiotic
drive), o más en general "conflicto intragenómico". Al igual que
la selección natural clásica, se trata de un proceso de
competencia, pero no entre individuos dentro de una especie, ni
entre especies dentro de un ecosistema, sino entre genes dentro
de un genoma, es decir, entre las partes de un mismo individuo.
Esto es posible porque cada individuo produce miles o millones
de gametos (óvulos o espermatozoides, según su sexo), cada uno
con una combinación distinta de genes. Y hay genes que sesgan a
su favor la producción de gametos, de modo que se aseguran su
presencia en más de la mitad de los espermatozoides o los
óvulos, que es lo que les correspondería por azar. Estos genes
son auténticas bombas evolutivas, porque pueden imponerse en una
población en pocas generaciones aun cuando no hagan nada
beneficioso para el individuo que los alberga. Los demás genes
se ven forzados a adaptarse para convivir en el mismo genoma que
ellos, y esto conduce a las poblaciones por caminos separados
aun cuando sus entornos sean similares. Esto es la evolución por
"conflicto intragenómico".
En el ejemplo mencionado antes de las dos subespecies de
Drosophila pseudoobscura, USA y Bogotá, el grupo de Allen Orr,
de la Universidad de Rochester, acaba de demostrar que un solo
gen (llamado overdrive) es responsable a la vez de la
esterilidad de los híbridos entre las dos subespecies, y de
causar su propia representación en los gametos por encima del
50% que le correspondería por azar. "Nuestros resultados",
afirma Orr, "indican que el conflicto intragenómico, una forma
de adaptación al ambiente genómico interno, es una fuerza
importante en la especiación".
Otro descubrimiento reciente es la importancia crucial de las
duplicaciones de genes en la evolución. Las duplicaciones o
pérdidas de genes son la principal fuente de variación genética
en nuestra especie: cualquier persona se distingue de cualquier
otra en un promedio de 70 regiones duplicadas o amputadas en uno
de sus cromosomas.
Dos siglos después, la ciencia rellena huecos que a Darwin le
hubiera encantado explicar.
Hoy se cumplen 200 años del
nacimiento del científico en Shrewsbury. La teoría de la
evolución resiste las envestidas creacionistas
El debate intelectual se produce en las universidades de Gran
Bretaña | La Iglesia defiende que creacionismo y evolucionismo
son complementarios | Las iglesias anglicana y católica han
sufrido un declive del 27% en asistencia a misa.
Doscientos años después de su
nacimiento, la teoría de Charles Darwin sobre la evolución de
las especies para explicar el origen de la vida goza de más
respeto que nunca entre científicos e intelectuales, y soporta
las embestidas periódicas de grupos religiosos –sobre todo en
Estados Unidos– empeñados en desprestigiarla para justificar sus
creencias y aumentar su base electoral e influencia política.
En una cuestión tan intrínseca a
la naturaleza humana, es inevitable que exista un intenso debate
que tradicionalmente ha enfrentado creacionismo y evolucionismo,
pero al que se ha incorporado de modo reciente –en una hábil
maniobra de organizaciones interesadas en difundir una
explicación religiosa o pseudorreligiosa del origen de la vida–
la llamada teoría del diseño inteligente.
Esta teoría no habla
estrictamente de un dios, sino de un ser superior con capacidad
para concebir y configurar el mundo tal y como lo conocemos.
Aunque la primera línea de fuego en esa guerra entre
evolucionistas y antievolucionistas está en los estados del
Medio Oeste y el cinturón industrial de EE.UU. (como Ohio, donde
el voto cristiano fue decisivo en las dos victorias electorales
de George Bush), el principal escenario del debate filosófico e
intelectual son las universidades y centros de estudios
científicos y teológicos de Inglaterra, una sociedad mucho más
laica que la norteamericana y donde el enfrentamiento no es ni
mucho menos tan venenoso.
"Podría decirse que lo que en Kansas o Pensilvania es casi un
combate sucio para que se enseñe el creacionismo en los colegios
públicos a pesar del amplísimo apoyo académico a las teorías de
Darwin, en el Reino Unido es un pulso entre caballeros de
acuerdo a las más estrictas normas del fair play donde ni una
parte ni la otra pierde los papeles", dice el profesor de
teología Leo Bentley. Un ejemplo es que la campaña humanista con
anuncios en el metro de Londres y autobuses de todo el país,
bajo el eslogan Dios probablemente no existe, así que no te
preocupes y disfruta, se ha desarrollado sin tan siquiera una
décima parte de la polémica suscitada en Barcelona.
Ello no quiere decir que no existan opiniones contrarias, como
demuestra una encuesta encargada por el think tank religioso
Theos con ocasión del 200.º aniversario del nacimiento de
Charles Darwin en Shrewsbury, que se celebra hoy, y 150.º
aniversario de la publicación de su obra maestra El origen de
las especies, efectuada por la firma de sondeos ComRes entre
poco más de 2.000 ciudadanos británicos. Los resultados son
difíciles de interpretar, por la manera en que se formularon las
preguntas y el hecho de que mucha gente diera credibilidad tanto
al creacionismocomo a la teoría del diseño inteligente, en vez
de escoger entre una y otra.
A la pregunta de "¿a qué atribuye usted el origen y desarrollo
de la vida?", un 48% de los encuestados respalda la teoría
darwiniana de la evolución, un 22% el creacionismo, un 17% la
teoría del diseño inteligente, y el resto dice que no sabe. En
cuanto a la pregunta de cuáles de estas explicaciones deberían
formar parte del expediente académico de los colegios públicos
de Gran Bretaña (con la posibilidad de optar por varias), un 69%
dice que el evolucionismo, un 44% que el creacionismo, y un 41%
que el diseño inteligente.
Además, un 51% está de acuerdo con la afirmación de que "la
evolución por sí misma no es suficiente para explicar las
complejas estructuras de algunos seres vivos, y por tanto debe
haber sido necesaria la intervención de un diseñador". El
problema es que la separación constitucional entre Iglesia y
Estado constituye un obstáculo para la difusión de cualquier fe
o precepto religioso en las escuelas estatales, y la creación
por Dios del mundo en siete días, y de Eva a partir de una
costilla de Adán, es uno de los pilares del cristianismo.
"Tenemos suerte de que, comparados con EE.UU. y otros países,
aquí no existe ningún grupo religioso o cultural de peso que se
oponga a la enseñanza de la teoría de la evolución", señala lord
Martin Rees, presidente de la Royal Society.
La cuestión en Gran Bretaña no es –como en algunos lugares de
EE.UU. bajo presión de la derecha religiosa que contribuyó a
auparaBush al poder– si el evolucionismo debería de dejarse de
enseñar en los colegios públicos, sino si debería acaso ir
acompañado del creacionismo o/y el diseño inteligente. A lo cual
dos tercios de los maestros responde que no, y un tercio que sí.
La difusión por 60 escuelas británicas de un DVD creacionista
procedente de Estados Unidos ha sembrado la alarma en el
gobierno del Labour, que ha exigido su retirada porque ni el
creacionismo ni el diseño inteligente son teorías científicas
admitidas, y –en palabras del diputado Graham Stringer-"bajo
ninguna circunstancia deben ser enseñados a la misma altura que
la teoría de la evolución". "La atribución del desarrollo de la
vida a una inteligencia superior es un sucedáneo religioso, una
manera de envolver de una manera más neutra el concepto básico
de que el mundo es obra de Dios", señala el teólogo Alfred
Borthwick.
La Iglesia de Inglaterra es la primera que se aleja de las
denuncias al darwinismo y quiere aprovechar el aniversario para
difundir la noción (igual que el Vaticano) de que evolucionismo
y creacionismo no son antagónicos sino complementarios, y la
teoría del origen y evolución de las especies es perfectamente
compatible con la intervención de Dios. "Dos siglos después de
tu nacimiento, te debemos disculpas por haberte malinterpretado
y animado a otros aque te malinterpreten", dice un mensaje
póstumo a Darwin colocado en la web de la Iglesia anglicana.
La ausencia de un movimiento antievolucionista en Gran Bretaña
se explica en el carácter cada vez más laico de la sociedad de
este país, donde nueve millones de habitantes (un 15% de la
población) son ateos o agnósticos, el grupo más importan después
de los cristianos (un 71,8%, pero la gran mayoría de tipo
social, es decir, que se definen como tales por tradición o
pertenencia a un grupo cultural, pero sin compartir
necesariamente los principios de la fe o tener una participación
activa en la religión).
Todas las encuestas sobre religiosidad están llenas de
contradicciones. Aunque un 75% de los habitantes del Reino Unido
se consideran cristianos, musulmanes, judíos, budistas o sijs,
al mismo tiempo un 66% afirma "no tener ninguna conexión
práctica con la religión o la Iglesia", y menos de la mitad dice
creer en Dios. Un sondeo realizado entre niños de nueve y diez
años del condado de Cornualles, el más pobre de Inglaterra,
concluye que sólo el 19% se ve a sí mismo como "religioso".
Desde 1980 hasta ahora, las Iglesias anglicana y católica han
sufrido un declive del 27% en asistencia a misa, y la única
iglesia institucional que ha registrado un aumento de fieles es
la ortodoxa griega y rusa, debido a la inmigración.
Para celebrar el 200.º aniversario del nacimiento de Darwin,
mañana se abre al público como museo la casa de Orpington (Kent)
donde el científico realizó sus experimentos y elaboró sus
teorías, con una detalla explicación de su vida, trabajo y
familia. El Museo de Historia Natural le dedica una exposición
con el relato del viaje del Beagle,sus escritos personales y la
ambientación de los invernaderos donde cultivaba sus orquídeas,
y se han organizado congresos en Cambridge, Shrewsbury y otras
ciudades.
La fe es algo que se tiene ó no
se tiene; no entiendo esas corrientes tan de moda que quieren
convencer a la gente de la existencia de Dios. Las creencias,
creo, son algo muy íntimo. En la Edad Media o "creías´" ó
"creías" si no querías morir en la hoguera. Galileo, según tengo
entendido, era una persona muy religiosa pero no por ello negaba
la evidencia: La Tierra no era el centro del Universo por mucho
que la Iglesia se empeñara. Menos mal que hoy en día y Ya en
tiempos de Darwin la Iglesia no tiene el poder de decidir lo que
es Verdad.
En cuanto al orígen del Ser
Humano, la evidencia es innegable por mucho que algunas
religiones quieran convencernos de lo contrario. No hay más
ciego que el que no quiere ver. La travesía del ser Humano ha
sido larga y dura.. muy dura; BASTA YA DE SUBESTIMARNOS.
Bibliografía Sugerida.
Castrodeza,
Carlos. Ortodoxia darwiniana y progreso biológico. Madrid:
Alianza Editorial, 1988. Obra de carácter divulgativo.
Darwin, Charles
R. Viaje de un naturalista alrededor del mundo. 2 tomos,
Madrid: Ediciones Akal, 1983. Viaje que realizó el autor
alrededor del mundo.
Darwin, Charles
R. Autobiografía. 2 vols., Madrid: Alianza Editorial, 1984.
Obra autobiográfica de Darwin.
Darwin, Charles
R. El viaje del Beagle. Barcelona: Editorial Labor, 1984.
Viaje que realizó el autor alrededor del mundo.
Darwin, Charles.
Origen de las especies. 2ª ed., 1994. Madrid: Ediciones Akal.
Libro donde el autor fundamenta su teoría de la evolución.
Ghiselin,
Michael T. El triunfo de Darwin. Madrid: Ediciones Cátedra,
1983. Obra divulgativa sobre la teoría de la evolución de
Darwin.
Howard,
Jonathan. Darwin. Madrid: Alianza Editorial, 1987. Obra
biográfica sobre la figura de Charles Darwin.
Mayr, Ernest.
Una larga controversia: Darwin y el darwinismo. Barcelona:
Editorial Crítica, 1992. Obra analítica sobre Darwin y sus
ideas.
Papp, Desiderio.
Darwin. La aventura de un espíritu. Madrid: Editorial Espasa-Calpe,
1983. Obra biográfica sobre la figura de Darwin.
Ruse, Michael.
La revolución darwinista. Madrid: Alianza Editorial, 1983.
Obra divulgativa sobre la teoría de la evolución de Darwin
Charles Darwin y su paso
por Argentina.
Por Graciela Cutuli. Tomado de Pagina 12.
El 2009 fue
declarado el Año Darwin, en homenaje al
naturalista inglés nacido hace 200 años. Durante
su viaje a bordo del “Beagle”, exploró las
costas argentinas, cuyo estudio contribuyó a su
teoría sobre el origen de las especies. Un
itinerario por los lugares que visitó el notable
científico.
Hace casi
170 años, un joven naturalista llamado
Charles Darwin zarpaba del puerto británico
de Plymouth a bordo de la nave “HMS Beagle”,
al mando del capitán Robert Fitz Roy. La
expedición, que duraría cinco años –desde
1831 a 1836– recorrió las costas de Africa,
Sudamérica y Australia, y permitió al
estudioso realizar las observaciones que
años más tarde desembocaron en su teoría
sobre la evolución y selección de las
especies. En su largo periplo pasó por
Luján, Bahía Blanca, Pehuen-Có, Puerto
Deseado y Tierra del Fuego, entre muchos
otros lugares donde observó, recogió
muestras y tomó notas y apuntes que luego se
convirtieron en el Diario de viaje de un
naturalista alrededor del mundo, uno de los
éxitos editoriales de su tiempo. Dos siglos
más tarde, entre algunos paisajes cambiados
y otros intactos, su extraordinario aporte
pone a nuestro país en el mapa del turismo
científico.
EL PUENTE QUE YA NO
ESTA Darwin llegó a la
desembocadura del Río Negro el 3 de agosto
de 1833, desde allí comenzaría largas y
detalladas incursiones en nuestro
territorio, buscando y recolectando fósiles
de mamíferos y otros animales. Pasa por la
actual Carmen de Patagones (“la villa es
llamada indistintamente El Carmen o
Patagones. Está edificada frente a un peñón
que mira al río y muchas de las casas han
sido excavadas en la arenisca”) en la
frontera con los territorios indios, visita
la salina cercana y avanza por el valle del
río Negro y el río Colorado. Se entrevista
con Juan Manuel de Rosas y se toma varias
líneas para describir la hermosura de las
jóvenes indias que cruza en el camino.
Atraviesa la zona de médanos del sur de la
provincia de Buenos Aires y llega a Bahía
Blanca, que sin duda no lo impresiona
demasiado: “Apenas merece el nombre de
aldea”, apunta. Pero las tierras de la
región, entre Bahía Blanca, la actual Pehuen-Có
y Punta Alta, sin duda estaban destinadas a
sorprenderlo, por la abundancia de fósiles y
restos de animales ya extinguidos. Hoy, el
museo de ciencias naturales de Punta Alta y
su dependencia, el museo Ameghino de Pehuen-Có,
conservan algunos restos de los megaterios,
macrauquenias y gliptodontes que
impresionaron a Darwin y lo llevaron a
investigar con detenimiento las formaciones
geológicas de la región, la fauna y la
flora. Desde Bahía Blanca, Darwin puso rumbo
a Sierra de la Ventana y Buenos Aires, que
describe como una ciudad grande y de trazado
regular, con calles en ángulo recto y casas
de un solo piso. Siguiendo viaje, cuenta:
“Hemos dejado atrás la pequeña ciudad de
Luján, donde hay un puente de madera sobre
el río, cosa rara en este país. También
hemos pasado por Areco. Las llanuras
parecían horizontales, es decir a perfecto
nivel, pero en realidad no era así, porque
en muchos sitios el horizonte estaba
distante. Aquí hay grandes extensiones
abandonadas entre estancia y estancia, pues
los buenos pastos escasean a causa de estar
la tierra cubierta de macizos de trébol acre
y cardos gigantes”, escribe en su diario.
Este puente, de 27 metros de largo y
levantado en madera de ñandubay, había sido
construido en 1773 por Pablo Márquez en
tierras de su propiedad, y provocó el
desplazamiento del Camino Real. Hoy ya no
existe, aunque el sitio es Lugar Histórico
Nacional y sus alrededores fueron escenario
de importantes hechos y acontecimientos en
la campaña libertadora. Sin embargo, Darwin
confiesa que el sitio no le importaba
demasiado, porque “apenas se hallan otros
cuadrúpedos y aves que la vizcacha y su
compañero ordinario, el mochuelo”.
|
 |
AL CORAZÓN DEL PARANÁ
El naturalista, fascinado con la
exuberancia de la naturaleza y
admirado de las costumbres de los
locales, sigue remontando el Paraná:
San Nicolás, Rosario, Santa Fe y río
arriba, hasta volver a Buenos Aires
y cruzar a Colonia por el Río de la
Plata, “una anchurosa extensión de
agua cenagosa sin grandiosidad ni
belleza”. Después de una excursión
por la Banda Oriental, vuelve a
zarpar de Buenos Aires rumbo a
Puerto Deseado, donde desembarca con
entusiasmo: “El primer desembarco en
un país nuevo es interesante y
especialmente cuando, como en este
caso, el aspecto del conjunto lleva
el sello de una individualidad bien
definida. A la altura de 60 a 90
metros sobre algunas masas de
pórfido, se extiende una vasta
llanura, que es peculiar y
característica de la Patagonia”. El
naturalista y sus acompañantes
remontan la ría Deseado y llegan
hasta el lugar que hoy se conoce
como “los miradores de Darwin”, un
paisaje espléndido y desolado, de
belleza sin fin: “El sitio en que
vivaqueamos estaba cercado de
atrevidos ricos y empinados
pináculos de pórfido.
No
creo haber visto nunca lugar más
apartado del resto del mundo que
esta gran grieta rocosa en la
extensa llanura”. Para los lectores
en lengua inglesa, es un lugar
mítico, que muchos quieren conocer
durante su paso por Puerto Deseado
en busca de descubrir la increíble
avifauna de la ría. Los miradores
pueden recorrerse desde lo alto,
llegando por tierra, o bien
navegando el cañón del río Deseado:
de un modo u otro, las vistas son
impresionantes. Darwin continuó
luego hacia el sur, hasta Puerto San
Julián, donde encontró un medio
esqueleto de Macrauquenia
patagónica, un cuadrúpedo
grande como un camello: “Es
imposible –escribe– reflexionar
sobre el cambio que se ha realizado
en el continente americano sin
sentir el más profundo asombro. En
remotas épocas, América debe haber
sido un hervidero de grandes
monstruos; ahora no hallamos más que
pigmeos, cuando se los compara con
las razas afines que los han
precedido”. |
|
>>>>>
Imágenes superiores: Los miradores
de Darwin, donde se unen la ría y el
río Deseado. Un horizonte sin fin
entre los macizos de pórfido. Islas
Malvinas. Uno de los puntos que tocó
el “Beagle” durante su largo viaje
de cinco años. Una foto de Charles
Darwin cuando tenía alrededor de 78
años. Huellas de megaterios, en la
costa de Pehuen-Có, donde Darwin
también descubrió una retama
endémica de la zona. |
Los expedicionarios aún
estaban lejos del fin del viaje. Siempre en
el extremo sur de la Patagonia, navegan el
río Santa Cruz, cuya descripción es idéntica
a la visión que podría tener el viajero de
hoy, con su “agua de un hermoso color azul,
pero con un ligero tinte lechoso. Fluye por
un lecho de guijarros parecido a los que
forman las riberas y llanos de los
alrededores. Describe una trayectoria
sinuosa por un valle que se extiende en
línea recta hacia el oeste. La anchura del
valle varía entre cinco y diez millas, y
está limitado por terrazas escalonadas, que
se levantan en casi todas partes, una sobre
otra, a la altura de 150 metros,
correspondiéndose de manera notable en las
orillas opuestas”. Una vez más, Darwin se
asombra de la uniformidad de fauna y flora
en la Patagonia, poblada por grandes manadas
de guanacos, multitud de pequeños roedores y
el puma, visible sobre todo a través de sus
huellas. El grupo avanza por el río Santa
Cruz hasta la cordillera, donde avista el
cerro Fitz Roy, así bautizado años más tarde
por el Perito Moreno en homenaje al capitán
del “Beagle”. En contraste con la desolación
que encontraron los expedicionarios, hoy el
Chaltén –pueblo jovencísimo nacido
oficialmente en 1986– es hoy una de las
capitales del turismo aventura en nuestro
país, impulsado por el espectacular
crecimiento del turismo en El Calafate. Sin
embargo, en época de Darwin y sus compañeros
la dureza de las condiciones para avanzar
los obliga a regresar hacia la costa
atlántica, para poner rumbo a las Islas
Malvinas –”un país ondulante, de aspecto
mísero y desolado, cubierto en todas partes
por un suelo turboso y una hierba fina y
dura, que presenta un color pardusco y
uniforme”– y, finalmente, tocar el último
punto en la actual Argentina: Tierra del
Fuego.
LA ISLA
La experiencia de Darwin al avistar
Tierra del Fuego es sin duda irrepetible
hoy: demasiada agua corrida bajo el puente y
la extinción de los nativos de la isla
permiten sólo imaginar lo que puede haber
sido su desembarco en ese brumoso territorio
del fin del mundo. Tras anclar en la Bahía
del Buen Suceso, describe la extraña
recepción de los indios, que se ocultaban en
el bosque lanzando un fuente clamor mientras
agitaban sus mantos, desde un pico que salía
sobre el mar. A la noche “vimos sus hogueras
y escuchamos de nuevo sus gritos salvajes”,
evoca, describiendo con minuciosidad sus
saludos, costumbres y hasta pequeñas
vanidades, como la del indígena que se
vanagloria de su altura y “se tiene por el
hombre más hermoso de Tierra del Fuego”. O
aquellos que por su total desconocimiento
del peligro de las armas de fuego no se
movían siquiera ante los disparos,
tomándolos tal vez por algún golpe u otro
fenómeno de poca letalidad.
Al naturalista le basta
además una ojeada para descubrir que todo lo
que lo rodea en la isla es muy distinto de
la Patagonia continental. A pesar de las
incomodidades, y en algunos casos del mal
tiempo, su descripción del Canal de Beagle
es entusiasta: “El paisaje aquí crece en
grandiosidad. Las altas montañas del lado
norte forman el eje granítico, o espinazo
del país, y se elevan súbitamente 900 o 1000
metros, culminando en un pico que sube a
unos 2000 metros. Están cubiertas de un
amplio manto de nieves perpetuas; numerosas
cascadas vierten sus aguas, por entre
boscaje, en el hondo canal angosto. En
muchas partes se extienden magníficos
glaciares desde la ladera de los montes
hasta el mar. Apenas es posible imaginar
algo más bello que el azul berilo de estos
glaciares, en especial por el contraste con
la blancura mate de la nieve que corona las
cimas”.
Como si los años no hubieran
pasado, como si el territorio salvaje de
ayer no se hubiera civilizado conservando
grandiosidad, en las orillas de Tierra del
Fuego, frente a las cumbres imponentes y los
colores contrastantes del mar austral y los
bosques, todo se puede mirar con los mismos
ojos de Darwin. Que siguió su viaje, rumbo a
las Galápagos y tantos otros lugares
fantásticos del globo aún casi sin explorar
en ese entonces, llevando en su diario y su
equipaje las semillas de avances
revolucionarios que cambiarían para siempre
la historia de la ciencia y la mirada del
mundo sobre su propio, lejanísimo pasado.
La selección natural; o
la supervivencia de los más aptos.
Fuente: Darwin, Charles. El origen
de las especies. Traducción de Juan Godo. Barcelona: Ediciones
Zeus, 1970. Fragmento de El origen de las especies. Capítulo IV.
Si, bajo
condiciones variables de vida, los seres orgánicos presentan
diferencias individuales en casi todas las partes de su
estructura, cosa que no puede discutirse; si hay una lucha
rigurosa por la existencia, debido a la proporción
geométrica de aumento en alguna época, estación o año, y
esto tampoco puede discutirse; considerando la infinita
complejidad en las relaciones de todos los seres orgánicos
entre sí y con sus condiciones de vida, origen de infinita
diversidad de estructura, constitución y hábitos que han de
ser ventajosos, sería un hecho muy extraordinario el que
nunca se hubiesen producido variaciones útiles para el
propio bienestar de cada ser, de la misma manera que se han
producido tantas variaciones útiles para el ser humano.
Mas si alguna
vez se producen variaciones útiles para cualquier ser
orgánico, seguramente los individuos así caracterizados
tendrán la mayor probabilidad de ser conservados en la lucha
por la vida; y debido al fuerte principio de la herencia,
tenderán a producir descendencia caracterizada de un modo
parecido. A este principio de conservación, o supervivencia
de los más aptos, yo le he dado el nombre de Selección
Natural. Conduce a la mejora de toda criatura en relación
con sus condiciones orgánicas e inorgánicas de vida; y por
consiguiente, en la mayoría de los casos, a lo que debe
considerarse como un progreso en la organización. Sin
embargo, las formas bajas y simples durarán mucho tiempo si
están bien adaptadas para sus condiciones de vida también
simples.
La selección
natural, basada en el principio de que las cualidades se
heredan en las edades correspondientes, puede modificar el
huevo, la semilla o la cría, tan fácilmente como al adulto.
Entre muchos animales, la selección sexual habrá prestado su
ayuda a la selección ordinaria, asegurando a los machos más
vigorosos y mejor adaptados el mayor número de
descendientes. La selección sexual dará también caracteres
útiles a los machos solos, en sus luchas o rivalidad con
otros machos; y estos caracteres serán transmitidos a un
solo sexo o a ambos sexos, según la forma de herencia que
predomine.
Si la selección
natural ha actuado realmente de este modo adaptando las
diversas formas de vida a sus diversas condiciones y
estaciones, debe juzgarse por el tenor general y el número
de las pruebas en pro y en contra que presentamos en los
capítulos siguientes. Pero ya hemos visto que ello acarrea
la extinción, y hasta qué punto la extinción ha actuado en
la historia del mundo, la geología nos lo indica claramente.
Asimismo, la selección natural conduce a la divergencia de
carácter; porque cuanto más diverjan los seres orgánicos en
estructura, hábitos y constitución, tanto más puede
sostenerse un número grande de individuos en la misma
región, de lo cual tenemos una prueba con sólo mirar a los
habitantes de cualquier lugar pequeño y a las producciones
naturalizadas en tierras extranjeras. Por lo tanto, durante
la modificación de los descendientes de cualquier especie, y
durante la lucha incesante de todas las especies por
aumentar en número, cuanto más diversificados lleguen a ser
los descendientes, mayores serán sus probabilidades de éxito
en la lucha por la vida. Así, las pequeñas diferencias que
distinguen a las variedades de la misma especie tienden
constantemente a aumentar, hasta que igualan las mayores
diferencias entre las especies del mismo género, o incluso
de géneros distintos.
Ya hemos visto
que la especie que más varía es la especie común, muy
difundida y muy distribuida, perteneciente a los géneros más
numerosos dentro de cada clase; y estas especies tienden a
transmitir a sus modificados descendientes aquella
superioridad que ahora las hace ser dominantes en sus
propios países. La selección natural, como acaba de
observarse, conduce a la divergencia de carácter y a una
gran extinción de las formas de vida menos perfeccionadas e
intermedias. A base de estos principios puede explicarse la
naturaleza de las afinidades y las distinciones generalmente
bien definidas entre los innumerables seres orgánicos dentro
de cada clase en el mundo entero. Es un hecho realmente
maravilloso (pero tendemos a no considerarlo maravilloso,
porque estamos familiarizados con el mismo) el de que todos
los animales y todas las plantas a través de todo el tiempo
y de todo el espacio se relacionen unos con otros formando
grupos, subordinados a grupos, tal como observamos en todas
partes, a saber, variedades de la misma especie
estrechamente relacionadas, formando secciones y subgéneros,
especies de distintos géneros mucho menos estrechamente
relacionados, y géneros relacionados en diferentes grados,
formando subfamilias, familias, órdenes, subclases y clases.
Los varios grupos subordinados dentro de cualquier clase no
pueden clasificarse en una sola fila, sino que parecen estar
arracimados alrededor de puntos, y éstos alrededor de otros
puntos, y así sucesivamente en ciclos casi interminables. Si
las especies hubiesen sido creadas independientemente, no
habría sido posible explicar esta clase de clasificación
pero se explica por la herencia y la acción compleja de la
selección natural, que provoca la extinción y la divergencia
de carácter, según hemos visto ilustrado en el diagrama.
Las afinidades
de todos los seres de la misma clase se han representado a
veces mediante un gran árbol. Creo que este símil dice en
gran parte la verdad. Las ramas verdes y florecientes pueden
representar las especies existentes; y las producidas
durante años anteriores pueden representar la larga sucesión
de especies extinguidas. En cada período de crecimiento,
todas las ramas, al crecer, han tratado de extenderse en
todos los sentidos y de superar y matar a las ramitas y
ramas que las rodeaban, de la misma manera que las especies
y los grupos de especies han vencido en todo tiempo a otras
especies en la gran lucha por la vida. Los troncos divididos
en grandes ramas y éstas en ramas más y más pequeñas, fueron
también en otro tiempo, cuando el árbol era joven, retoños
florecientes; y esta conexión de los brotes antiguos y
actuales en los ramificados brazos puede representar a las
mil maravillas la clasificación de todas las especies
extinguidas y vivientes en grupos subordinados a otros
grupos. De los muchos retoños que florecieron cuando el
árbol no era más que un arbusto, sólo dos o tres,
convertidos en grandes ramas, sobreviven todavía y sostienen
las otras ramas; lo mismo ocurre con las especies que
vivieron durante los remotos períodos geológicos, las cuales
muy pocas han dejado en pos de sí descendientes vivientes y
modificados. Desde el primer crecimiento del árbol, más de
una rama de todos los tamaños se ha deteriorado y caído; y
estas ramas caídas de diversos tamaños pueden representar
aquellos órdenes, familias y géneros enteros que ahora no
tienen representantes vivientes y que sólo nos son conocidos
en su estado fósil. De la misma manera que aquí y allá vemos
una ramita solitaria que sale de la parte baja del tronco de
un árbol, que por alguna circunstancia fortuita ha sido
favorecida y todavía está viva en su parte superior, así
vemos ocasionalmente un animal como el ornitorrinco o el
lepidosiren, que en grado exiguo enlaza por sus afinidades a
dos grandes ramas de la vida y que, al parecer, se ha
salvado de la fatal competencia por haber habitado en un
paraje resguardado. Del mismo modo que los retoños dan
origen, al crecer, a otros retoños, y éstos, cuando son
vigorosos, se ramifican y dominan por todos lados a muchas
ramas más débiles, así creo que ha sucedido, por medio de la
generación, con el gran Árbol de la Vida, que llena la
corteza de la Tierra con sus ramas muertas y rotas, y cubre
la superficie con sus incesantes y hermosas ramificaciones.
|
 |
En 2009 se celebrará en todo el mundo el 150
aniversario de la publicación de dicha obra y
200 del nacimiento de su autor. Pero, más allá
de los festejos, será un año para reflexionar
sobre su legado, repensarlo, valorarlo,
difundirlo y trasladarlo a nuestro tiempo, en el
cual sus teorías recobran vigencia de manera
continua. El
24 de noviembre se cumplen 150
años de la publicación del libro de Charles
Darwin, por tal motivo, la Asociación Amigos del Museo Municipal Punta Hermengo de Miramar
y Grupo Paleo Contenidos, presenta a modo de
homenaje una interesante publicación titulada:
Un Siglo y Medio de “El Origen de las Especies”
de Chales Darwin.
Mariano Magnussen Saffer,
presidente de la Asociación Amigos del Museo
Municipal Punta Hermengo y autor de la
publicación, recorre en 166 paginas la vida de Charles
Darwin, desde su juventud hasta el momento de
escribir su obra, y una traducción y adaptación perfecta de
la obra.
<<<
Obtener Publicación en Pdf
Gratis!!!.
|
 Ver
mas Artículos de Divulgación Paleontológica Internacional
Ver
mas Artículos de Divulgación Paleontológica Internacional |