Por Mariano
Magnussen Saffer. Grupo Paleo. Museo Municipal Punta Hermengo de
Miramar. Fundación Argentavis.
marianomagnussen@yahoo.com.ar. Tomado de - Magnussen Saffer,
Mariano. 2016.
El origen de los caballos y su proceso evolutivo. Paleo Revista Argentina de
Paleontología. Boletín Paleontológico. Ańo XIV. 140: 15-21.
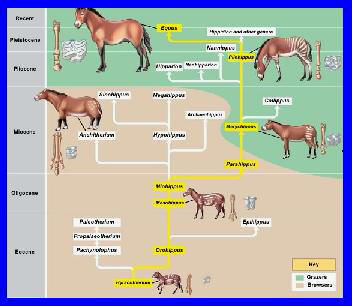
Los primeros
ungulados aparecen en Asia en la época del Paleoceno superior,
dentro de la Era Cenozoica o Terciaria. El caballo pertenece a la
orden del perisodáctilos, es decir, los ungulados provistos de un
número impar de dedos. Uno de estos dedos es predominante y asegura
el principal apoyo del animal.
El registro fósil
nos permite contemplar los cambios evolutivos a través de la mayor
escala temporal. Podemos ver a las especies aparecer y extinguirse
repetidamente a lo largo del registro fósil. Una especie animal
puede, por termino medio, sobrevivir entre uno a diez millones de
ańos, aunque con muchas variaciones.
Al
estudiar la sustitución de unas especies por otras especies
u otros taxones a lo largo del tiempo, se observan
tendencias.
Las
tendencias son cambios orientados en los rasgos
característicos o en los modelos de diversidad en un grupo
de organismos. Las tendencias fósiles demuestran claramente
el principio de Darwin del cambio perpetuo.
Hay que
hacer hincapié en que las tendencias se observan solamente a
posteriori. No podemos predecir, a partir de los primeros
fósiles de un grupo, cual será el aspecto o la diversidad de
los fósiles posteriores. El proceso evolutivo no contiene
direcciones predeterminadas.
Los
primeros équidos que existieron poseían cuatro dedos en
cada extremidad de sus miembros anteriores y tan sólo tres
extremidades en los posteriores. Como resultado de la evolución,
para una adaptación progresiva a la carrera, el número de dedos que
descansan en el suelo, a lo largo de las edades, fue reduciéndose a
tres, posteriormente a dos, hasta la aparición del casco único,
característica del caballo actual.
Durante mucho
tiempo se ha creído que el antepasado más lejano del caballo era el
"Hyracotherium leporinum", aparecido durante el Eoceno, (hace
aproximadamente 54 millones de ańos) Pero, estudios recientes han
emplazado a los caballos entre los Palaeotherium.
Palaeotherium. Cuvier, 1804.
es
un género de mamíferos perisodáctilos de la familia Palaeotheriidae.
Medían cerca de 75 cm de altura, con rasgos parecidos al actual
tapir. Vivieron en los densos bosques tropicales de Europa hace
alrededor de 45 millones de ańos durante la primera mitad del
Eoceno.
George
Cuvier lo describió originalmente como una especie de tapir,
y como estos, Palaeotherium ha sido recosntruido como
un animal parecido a un tapir. El examen de los cráneos son
embargo ha mostrado que la cavidad nasal no era apta para
tener una trompa pequeńa, lo que ha derivado en nuevas
reconstrucciones que lo muestran como una criatura más
parecida a los caballos. Los estudios anatómicos también
sugieren que Palaeotherium, junto con otros géneros
de paleotéridos como Hyracotherium, estaban
cercanamente relacionados con los caballos.
En
general las especies de Palaeotherium medían cerca de 75
centímetros de altura hasta los hombros, pero la especie de mayor
tamańo, P. magnum del Eoceno medio de Francia, alcanzaba una
talla comparable a los mayores caballos modernos.
Eohippus.Marsh, 1876.
Es un género
extinto de mamífero ungulado pequeńo de la familia de los équidos.
La única especie reconocida es E. angustidens, el cual
tradicionalmente fue considerado como una especie del género
Hyracotherium. Sus restos se han hallado en América del Norte.
En 1876,
Othniel C. Marsh describió un esqueleto como Eohippus
validus, a partir del griego ηώς (eōs, "amanecer") y
ιππος (hippos, "caballo"), lo que significaría "caballo del
amanecer". Sus similitudes con los fósiles descritos por
Richard Owen fueron seńaladas formalmente en un artículo de
1932 de sir Clive Forster Cooper. E. validus fue
reclasificado en el género Hyracotherium, el cual
tenía prioridad como nombre de género, así que Eohippus
se volvió el sinónimo menor de este género.
Más recientemente
se encontró que Hyracotherium es un grupo parafilético de
especies, y ahora este género solo incluye a la especie H.
leporinum. Por su parte, se determinó que E. validus era
idéntico a una especie nombrada antes, Hyracotherium angustidens
(Cope, 1875), y el nombre binomial resultante es entonces
Eohippus angustidens.
Mesohippus.
Marsh, 1875.
Es un género
extinto de mamíferos perisodáctilos de la familia Equidae que
vivieron hace aproximadamente 40 millones de ańos, en el Oligoceno
Medio. Tenía las patas más largas que su predecesor Hyracotherium.
Perdió, también con respecto a éste, un dedo, aunque se apoyaba
normalmente sobre su dedo medio, a pesar que los otros dos también
eran usados.
La
cabeza de Mesohippus era más grande y larga que la de
sus ancestros. Sus ojos eran redondeados y dispuestos más
separados a los lados de la cabeza. Los dientes eran
igualmente más largos y parecidos a los de los caballos
actuales, al igual que su cavidad cerebral. Como la mayoría
de los caballos fósiles, Mesohippus era común en
Norteamérica.
Miohippus.
Marsh, 1874.
Es un género extinto de
équidos de la subfamilia Anchitheriinae. Los fósiles se encuentran en
Norteamérica; parece haber evolucionado del género Mesohippus,
con el que coexistió durante cuatro a ocho millones de ańos.
Los
ejemplares pesarían entre 40 y 55 Kg, siendo más grandes que la
mayoría de los primeros équidos del Eoceno, pero menores que los
caballos actuales. Era algo más grande que Mesohippus, y
con el cráneo ligeramente más largo. También tenía una cresta
adicional en sus molares superiores, lo que le permitía masticar
hierba con mayor efectividad. Algunas especies estaban adaptadas
a la vida en los bosques, y otras a la vida en las praderas.
Miohippus
fue nombrado por Marsh en 1874 y su especie tipo es Miohippus
annectens. Fue asignado a la familia Equidae por Marsh en 1874. Fue
sinonimizado subjetivamente con Mesohippus por Matthew en 1899.
Su estatus como género independiente fue corroborado por Hay (1902), H.
F. Osborn en 1918, Hay (1930), Stirton (1940) y Carroll (1988); y fue
asignado a la subfamilia Anchitheriinae por MacFadden en 1998.
Es un género extinto de
équido que vivió entre los 17 a 11 millones de ańos atrás en la época
del Mioceno. Su nombre significa “caballo rumiante” pero no se sabe si
en verdad rumiaba.
Los
Merychippus vivían en manadas, tenía una altura de 1,20 metros, tuvo
tres dedos en cada pie. Tenía el hocico más largo, la mandíbula más
profunda, los ojos más amplios que cualquier caballo hasta la fecha. Su
cerebro también fue mucho mayor haciéndolo más inteligente y ágil y fue
el primero en tener la cabeza distintiva de los caballos actuales.
En algunas
especies de Merychippus, los dedos laterales eran más
largos, mientras que en otros, los tenían pequeńos y sólo tenían
contacto con el suelo cuando corrían.Sus dientes eran como los
de Parahippus, con la cresta extra que era variable en
Miohippus, se volvió permanente en Merychippuss, y en
los demás dientes se empiezan a formar unas series de crestas
altas con mayores coronas. Merychippus dio origen al
menos a 19 especies distintas pastadoras.
El
primer grupo lo constituyen las especies de Hipparion son
pastadores de 3 dedos y se dividen en 4 géneros y 16 especies. El último
fue una línea de “equinos verdaderos” que conduce a las especies
actuales, en donde sus dedos laterales eran más pequeńos que las de
otros proto-caballos.
Pliohippus.Marsh, 1874.
Es un
género extinto de mamíferos perisodáctilos de la familia de los équidos,
que vivió durante el Plioceno en América del Norte. Era herbívoro,
cuadrúpedo y de cuello largo y arqueado.
Surgió
de Calippus en el Mioceno Medio, alrededor de 12 millones de ańos
atrás. Era muy similar en apariencia al Equus, a pesar de que
tenía dos dedos largos en ambos lados de la pezuńa.
Hasta hace
poco, se creía que Pliohippus era el ancestro de los
actuales caballos, porque presentaba muchas de sus semejanzas
anatómicas. Sin embargo, aunque claramente Pliohippus fue
un pariente cercano del Equus, su cráneo presenta
profundas fosas faciales, mientras que en el Equus no
están esas fosas en absoluto. Además, sus dientes eran muy
curvos, a diferencia de los dientes bien rectos de los caballos
modernos.
En
consecuencia, es poco probable que sea el ancestro del caballo moderno,
en cambio, es un probable candidato para ser el antepasado de
Astrohippus.
Hipiddion.
Roth, 1899.
Llego a Sudamérica a
principios del Pleistoceno en el gran intercambio faunistico de fines
del Plioceno. De acuerdo a la recientes teorías de algunos
Paleontólogos, este genero seria originario de Sudamérica y estaría
emparentado con Pliohippus y Dinohippus del Plioceno de
América del norte. Era un primitivo caballo, el cual se extinguió poco
antes de la llegada de los primeros colonos sin dejar representantes
vivientes, siendo reemplazado por el caballo europeo. Era similar al
cebras de Africa, pero algo mas bajo y rechoncho, con extremidades
cortas.
Su cráneo
presenta unos huesos nasales algo alargados y pronunciados que
las formas vivientes. Por otro lado, es notable la conformación
de las extremidades, proporcionalmente cortas y anchas, que le
confieren al animal un aspecto macizo. Su peso pudo
ser de 400 kilos. El primer registro de un caballo fósil
sudamericano fue el molar superior descubierto por Charles
Darwin cerca de Bahía Blanca (Prov. Buenos Aires), al que
Richard Owen identifico en 1840, lo denomino Equus caballus
(la especie actual), y mas tarde lo denomino Equus curvidens.
Se han hallado numerosos esqueletos bien preservados en
distintas zonas del país.
El registro es
ampliamente conocido desde el norte de Ecuador hasta el sur Patagonico.
Desde el punto de vista estratigráfico, es conocido desde el Plioceno
superior (Uquiense) hasta el Pleistoceno superior - Holoceno (Lujanense
reciente). Las características adaptativas de la parte distal de las
extremidades de Hippidion pueden ser indicadores de distintos
tipos de suelo y vegetación.
El hábitat de
Hippidion debería corresponder a un bosque húmedo y suelo blando.
Los Equidos de América del sur se han extinguido totalmente. Algunos
ejemplares mas modernos de Hippidion, provienen de sitios
arqueológicos de la Provincia de Buenos Aires, como Arroyo Seco, ubicado
en el Partido de Tres Arroyos, con una antigüedad de 8500 ańos.
Equus.Linnaeus, 1758.
Se cree que durante el
Pleistoceno, hace unos 15.000 ańos, el género Equus extendió su
área de distribución desde Norteamérica a Eurasia y África cruzando el
puente de Beringia.
Hace unos
10.000 ańos los caballos se extinguieron en Norteamérica, por
causas aún desconocidas, quizá por algún cataclismo climático
que modificó los ecosistemas americanos. Diversos hallazgos en
cuevas de Europa indican que el caballo era un animal muy
abundante durante la Edad de Piedra en dicho continente; se han
encontrado suficientes restos de esqueletos de caballos dentro y
en los alrededores de estas cuevas como para afirmar que eran
consumidos por el ser humano.
El
número de caballos disminuyó en el neolítico, cuando Europa estaba
cubierta por bosques en su mayor parte. Se han encontrado restos de la
Edad del Bronce, embocaduras y piezas de arneses, que demuestran que el
caballo ya estaba domesticado en esta época. Esta domesticación siglos
más tarde permitió, tras el descubrimiento de América, que los caballos
fueran reintroducidos por los conquistadores espańoles en el continente
que los vio surgir.
Amerhippus.Hoffstetter, 1950
es un subgénero del
género Equus el cual agrupa a las 5 especies de dicho género de
la familia Equidae, que vivieron en el Pleistoceno medio al Holoceno
temprano de América del Sur, todas ellas se han extinguido. Los tamańos
de estos mamíferos perisodáctilos, si bien eran robustos y bastante
grandes, eran inferiores al de un caballo doméstico actual. Los
materiales de mayor antigüedad provienen del Ensenadense de Tarija, y
fueron fechados radiocarbónicamente con una edad de entre 1 millón y
700 mil ańos.
se
caracteriza por presentar una mandíbula robusta, y sus miembros
monodáctilos macizos y cortos. El cráneo exibe una cresta supraoccipital
ancha, y una flexión craneal destacada.
Su original
creador, Robert Hoffstetter, seńaló que Equus (Amerhippus)
Se distingue principalmente de Equus (Equus) por la
ausencia completa de la cavidad en forma de cono comprimido
—llamado cartucho externo o cornete— situada en el extremo libre
de los incisivos inferiores.
Esto sería el
resultado de una evolución regresiva, y trae como resultado la
pérdida de esmalte en la superficie que aplica masticación. Dos
ańos más tarde agregó que el cúbito es más fuerte en
Amerhippus que en otros subgéneros de Equus.
Otros autores
encontraron adecuado separar en un subgénero propio a los
Equus sudamericanos pues todas sus especies poseen unas
características extremidades más cortas y robustas, un cráneo
más grande (en relación a las proporciones corporales), y la
misma morfología dentaria.
Seguramente habitaban en espacios abiertos de estepas, praderas, o
sabanas, en pequeńos grupos que pastarían siempre atentos al peligro que
representaban los variados predadores carnívoros. Su dieta era
herbívora.
Sus
especies vivieron hasta el final del Pleistoceno o el Holoceno temprano,
por lo que convivieron durante algunos milenios con las primeras oleadas
humanas llegadas a América del Sur, es decir los primitivos amerindios.
Estos, según los especialistas, ejercieron una presión cazadora que
podría haber afectado el equilibrio poblacional, lo que podría ser una
de las causas de su completa extinción.
Equus caballus.Linneo, 1758
El caballo, vuelve a
América en los barcos de las “conquistas” espańolas en el siglo XV y XVI.
Por ejemplo en la primera expedición de Colón, quien arribó a las
Antillas y Bahamas, trajo 30 caballos, que dejó en Santo Domingo. En 10
ańos, la cantidad de caballos en esas regiones era muy importante.
Más tarde y de la misma
forma, llegan más caballos a México y Sudamérica. En tan sólo 400 ańos,
la población de caballos en América, alcanzo aproximadamente los 25
millones de cabezas siendo un tercio de la población equina del resto
del mundo.
A pesar de una
prohibición temporal que dictó Fernando el Católico, siempre se
siguieron enviando caballos a América. Dos ejemplos que pueden citarse
son los casos de don Pedro de Mendoza. que los trajo al Río de la Plata,
y Cabeza de Vaca que los llevó al Paraguay. Pero no todos los animales
embarcados llegaban a destino, porque siempre morían algunos por falta
de alimento y de cuidados o, sencillamente, porque cuando el agua
escaseaba los tiraban al mar.
No obstante estas
pérdidas, o las que se produjeron en las luchas con los nativos, o
motivadas por otras causas, como la ingestión de pastos tóxicos o por
picaduras de víboras venenosas, el equino se reprodujo fácilmente en
América debido a sus condiciones climáticas y a su vegetación.
Bibliografía sugerida.
Alberdi, María Teresa; Prado, José Luis (2004). «Caballos
fósiles de América del Sur: Una historia de tres millones de ańos».
INCUAPA, facultad de Ciencias Sociales, Universidad Nacional del Centro
de la Provincia de Buenos Aires (Olavarría, Argentina): 269
Froehlich, D. J. (2002). «Quo vadis eohippus? The systematics and
taxonomy of the early Eocene equids (Perissodactyla)». Zoological
Journal of the Linnean Society 134 (2): 141–256.
Joomun,
S. C.; Hooker, J. J.; Collinson, M. E. (2008). «Dental wear variation
and implications for diet: An example from Eocene perissodactyls (Mammalia)».
Palaeogeography, Palaeoclimatology, Palaeoecology 263 (3–4): 92.
MacFadden, B. J. (1984). "Astrohippus and Dinohippus". Journal of
Vertebrate Paleontology. 4 (2):273-283.
McKenna,
M. C.; Bell, S. K. (1997). Classification of Mammals: Above the Species
Level. Columbia University Press. p. 631.
MacFadden. F. 1998. Equidae. In C. M. Janis, K. M. Scott, and L. L.
Jacobs (eds.), Evolution of Tertiary Mammals of North America 1:537-559.
Orlando,
L.; et al. (2008). «Ancient DNA Clarifies the Evolutionary History of
American Late Pleistocene Equids». Journal of Molecular Evolution 66:
533–538.
Zacarías,
G.G.; Mińo-‐Boilini, A.R.; Zurita, A.E. & Francia, A. (2011). Campamento Vespucio, una
nueva localidad con mamíferos pleistocenos en la provincia de Salta,
Argentina. Estudios Geológicos [en línea], manuscrito aceptado, doi:
10.3989/egeol.40472.147.
La paleontología durante el Siglo XVII y la edad
de la razón.
Por Mariano Magnussen Saffer. Grupo
Paleo. Museo Municipal Punta Hermengo de Miramar. Fundación Argentavis.
marianomagnussen@yahoo.com.ar- Magnussen Saffer,
Mariano. 2016. La
paleontología durante el Siglo XVII y la edad de la razón.
Paleo Revista Argentina de
Paleontología. Boletín Paleontológico. Ańo XIV. 137: 11-13.
Durante
la Edad de la Razón, los cambios fundamentales en la filosofía natural
se reflejaron en el análisis de los fósiles. En 1665 Athanasius Kircher
atribuyó el origen de los huesos gigantes a razas extintas de humanos
gigantes en su Mundus subterraneus.
En el
mismo ańo, Robert Hooke publicó Micrographia, una colección
ilustrada de sus observaciones al microscopio. Una de estas
observaciones se titulaba "De madera petrificada, y otros organismos
petrificados", que incluía una comparación entre la madera petrificada y
la madera ordinaria. Concluyó que la madera petrificada era madera
ordinaria que había sido empapada con "agua impregnada con piedras y
partículas de tierra".
Sugirió
entonces que varios tipos de fósiles de conchas marinas se
habían formado a partir de conchas ordinarias mediante un
proceso similar.
Argumentó
en contra de la opinión generalizada de que tales objetos eran
"piedras formadas por alguna extraordinaria virtud plástica
latente en la Tierra misma". Hooke creía que los fósiles
proporcionaban evidencia sobre la historia de la vida en la
Tierra, como lo declaró en 1668:
... si
el hallazgo de monedas, medallas, urnas, y otros monumentos de
poblaciones o personas famosas, o sus utensilios se admiten como pruebas
incuestionables de que tales personas o cosas tienen, o en otros tiempos
tuvieron un ser; sin duda a estas petrificaciones se les permite tener
la misma validez y prueba de que antes han sido dichos vegetales o
animales... y son verdaderos personajes universales legibles para todos
los hombres racionales.
Hooke
estaba dispuesto a aceptar la posibilidad de que algunos de estos
fósiles representaban especies que se habían extinguido, posiblemente en
catástrofes geológicas pasadas.
En 1667
Nicolás Steno escribió un artículo acerca de una cabeza de tiburón que
había diseccionado. Comparó los dientes de tiburón con los objetos
fósiles comúnmente conocidos como piedras de lengua. Llegó a la
conclusión de que estos fósiles deberían haber sido los dientes de un
tiburón. Steno se interesó entonces en la cuestión de los fósiles, y
para hacer frente a algunas de las objeciones sobre su origen orgánico
comenzó a estudiar los estratos de suelo. El resultado de este trabajo
fue publicado en 1669 como Un precursor de una disertación acerca de
un sólido encerrado de forma natural dentro de otro sólido.
En este
libro, Steno estableció una distinción clara entre objetos como los
cristales de roca que realmente se habían formado dentro de las rocas y
las conchas fósiles o dientes de tiburón que se habían formado fuera de
esas rocas. Steno se dio cuenta de que ciertos tipos de rocas se habían
formado por la deposición sucesiva de capas horizontales de sedimento y
que los fósiles eran los restos de organismos vivos que se habían
enterrado en ese sedimento.
Como la mayoría
de los filósofos naturales del siglo
xvii. Steno creía
que la Tierra tenía solo unos pocos miles de ańos de antigüedad,
por lo que recurrió a la inundación bíblica como una posible
explicación para los fósiles de organismos marinos lejos del
mar.
A pesar de la
considerable influencia del Precursor, naturalistas como
Martin Lister (1638-1712) y John Ray (1627-1705) siguieron
cuestionando el origen orgánico de algunos fósiles. Ellos
estaban especialmente preocupados por objetos como los ammonites
fósiles, los cuales Hooke afirmaba eran de origen orgánico, sin
embargo no se parecían a ninguna especie viva conocida. Esto
planteaba la posibilidad de una extinción, que encontraron
difícil de aceptar por razones filosóficas y teológicas.
En 1695
Ray le escribió al naturalista galés Edward Lluyd quejándose de tales
puntos de vista: "...viene una serie de consecuencias tales, como las
que parecen escandalizar la Escritura-Historia de la edad del Mundo; al
menos derrocan la opinión vigente, y no sin razón, entre teólogos y
filósofos, que desde la primera Creación no ha habido especies de
animales o vegetales perdidas, ni han surgido nuevas".
Bibliografía
sugerida.
Bowler, Peter J. (2003).
Evolution:The History of an Idea. University of
California Press.
Bowler,
Peter J.; Iwan Rhys Morus (2005). Making Modern Science. The
University of Chicago Press.
Cadbury, Deborah
(2000), The Dinosaur Hunters: A True Story of Scientific Rivalry and
the Discovery of the Prehistoric World, Fourth Estate,
Desmond,
Adrian (1975). "The Discovery of Marine Transgressions and the
Explanation of Fossils in Antiquity". American Journal of Science,
Volume 275.
Palmer,
Douglas (2005) Earth Time: Exploring the Deep Past from Victorian
England to the Grand Canyon. Wiley, Chichester. I
Rudwick,
Martin J.S. (1985). The Great Devonian Controversy: The Shaping of
Scientific Knowledge among Gentlemanly Specialists. The University of
Chicago Press.
Zittel,
Karl Alfred von (1901). History of geology and palaentology to the
end of the Nineteenth Century. Charles Scribner's Sons, London.
Wikipedia.
Historia de la evolución de los Perisodáctilos.
Por Mariano Magnussen Saffer. Grupo Paleo. Museo
Municipal Punta Hermengo de Miramar. Fundación Argentavis.
marianomagnussen@yahoo.com.ar. Tomado de; Magnussen Saffer,
Mariano. 2016. Historia de la
evolución de los Perisodáctilos. Paleo Revista Argentina de
Paleontología. Boletín Paleontológico. Ańo XIV. 144: 15-17.
Los
Perisodáctilos son mamíferos ungulados que se caracterizan por la
posesión de extremidades con un número impar de dedos terminados en
pezuńas, y con el dedo central, que sirve de apoyo, más desarrollado que
los demás. Son herbívoros.
Los
perisodáctilos más primitivos se habían separado muy poco de los
condilartros. Hyracotherium, del Eoceno tenía el tamańo de un
perro pequeńo y se parecía al condilartro Phenacodus; tenía una
dentición completa, con premolares trituberculados y molares cuadrados
bunodontos con dos crestas transversales. La locomoción era digitígrada
en las formas primitivas.
Los
primeros perisodáctilos conocidos datan del Eoceno, aunque posiblemente
surgieron en Asia durante el Paleoceno Superior, menos de 10 millones de
ańos después de la extinción masiva del Cretácico-Terciario en la cual
se extinguieron los dinosaurios y otros muchos organismos. A principios
de Eoceno, hace 55 millones de ańos, ya estaban diversificados y
ocupaban varios continentes.
Los caballos y
tapires evolucionaron en Norte América mientras que los
rinocerontes parecen haber evolucionado en Asia a partir de
animales similares al tapir y luego haber recolonizado América
durante el Eoceno Medio (hace unos 45 millones de ańos).
Existieron 15 o 16 familias, de las cuales solo tres sobreviven
en la actualidad.
Dichas
familias fueron muy diversas en apariencia y tamańo; algunas incluían
animales gigantescos (Brontotheriidae) y extrańos (Chalicotheriidae). El
mayor perisodáctilo fue un rinoceronte asiático (Paraceratherium)
que, con 11 toneladas, fue más de dos veces mayor que los elefantes
actuales.
Los
perisodáctilos fueron el grupo de grandes mamíferos herbívoros dominante
durante el Oligoceno. No obstante, la expansión de las praderas durante
el Mioceno (hace unos 20 millones de ańos) favorecieron a los
artiodáctilos que, con sus estómagos provistos de cámaras se adaptaron
mejor a una dieta tan pobre en nutrientes y pronto les arrebataron la
supremacía. A pesar de ello, muchas especies de perisodáctilos
sobrevivieron y prosperaron hasta el final del Pleistoceno (hace sólo
10.000 ańos), cuando no pudieron soportar la presión de los cazadores
humanos ni el nuevo hábitat.
Los
primeros perisodáctilos eran muy similares a los otros ungulados
primitivos. Las patas desarrollan la condición mesaxónica, es decir, el
dedo central (tercero) se hace más grande y los demás se reducen, al
mismo tiempo que la región inferior de las extremidades se alarga y la
superior se acorta, con reducción de la ulna (cúbito) y la fíbula
(peroné).
Un rasgo
distintivo de los perisodáctilos es la organización del carpo y
del tarso. En las patas anteriores (a veces referidas como "manos"
en el habla popular), un carpiano distal, el hueso grande, se
ensancha y se articula con los carpianos proximales, mientras
que en el pie, el ectocuneiforme se transforma en un gran hueso
plano que transmite el empuje del astrágalo a través de un
navicular aplanado; el astrágalo tiene la superficie inferior
aplanada y no a modo de tróclea como en los artiodáctilos.
Entre
los primeros y mas pequeńos, encontramos a Hyracotherium, un género
extinto de ungulado perisodáctilo que fue hallado en la Formación
Arcilla de Londres en el Reino Unido. Este animal del tamańo de un perro
fue alguna vez considerado como el miembro más antiguo de la familia
Equidae antes de que su especie tipo, H. leporinum, fuera
reclasificada como un paleotérido, un tipo de perisodáctilos
relacionados tanto con los caballos como con los brontotéridos.
Hyracotherium
medía generalmente unos 60 centímetros de longitud y unos 20 centímetros
de altura hasta los hombros y pesaría cerca de 20 kilos. Tenía cuatro
dedos con pezuńa en cada pata delantera y tres en cada pata posterior.
Cada dedo poseía una almohadilla en su envés, similares a las de los
perros. Tenía un rostro corto con las cuencas oculares situadas a mitas
del cráneo y un corto diastema (el espacio entre los dientes frontales y
los de la mejilla).
El
cráneo entero era largo, con unos 44 dientes de corona baja. Aunque
fueran de corona baja, se puede ver que ya tenían los inicios de los
característicos molares con bordes de los caballos modernos.
Se cree que
Hyracotherium era un herbívoro ramoneador que comía
principalmente hojas suaves y también frutas y brotes de las
plantas.
Por otro lado,
en su máximo esplendor, fue Paraceratherium, un género
extinto de mamíferos perisodáctilos, suborden Ceratomorpha de la
familia Hyracodontidae. Se han descrito cinco especies
diferentes, pero tanto las relaciones entre dichas especies como
las relaciones a nivel de género han generado una controversia
que aún dura actualmente, un siglo después del descubrimiento de
los primeros fósiles.
Serían
los mamíferos terrestres mayores de todos los tiempos, con una altura de
cinco metros y medio en la cruz y de siete metros y medio en la cabeza,
y ocho metros de longitud (sin contar la cola). Las estimaciones del
peso de los paraceraterios varían, pero oscilan en torno a las quince
toneladas.
Los
paraceraterios, al ser ceratomorfos, están relacionados con los
rinocerontes actuales. Sin embargo, a diferencia del rinoceronte blanco,
se alimentaban arrancando las hojas de los árboles. Tampoco tenían
cuernos. Los animales de este género aparecieron en Asia central durante
la primera mitad del periodo Oligoceno, hace aproximadamente treinta
millones de ańos, y se extinguieron a mediados del periodo Mioceno, hace
16,6 millones de ańos.
Bibliografía sugerida.
Forster Cooper, C. (1934). The
extinct rhinoceroses of Baluchistan. Philosophical Transactions of the
Royal Society. 223, pp. 569-616. p. 48.
Hooker,
J.J. 2005. Perissodactyla; in K. D. Rose & J. D. Archibald (eds.), pp.
199-214: The Rise of Placental Mammals, Origins and Relationships of the
Major Extant Clades. The Johns Hopkins University Press, Baltimore.
J. J.
Hooker and D. Dashzeveg: The origin of chalicotheres (Perissodactyla,
Mammalia). en: Palaeontology: Vol. 47 Part 6, 2004, S. 1363–1386
Lucas, S. G.; Sobus, J. C. (1989). The
Systematics of Indricotheres (dentro de The Evolution of Perissodactyls).
Prothero, D. (Oxford University Press). Oxford University Press. p. 537.
McKenna,
M. C. & Bell, S. K. 1997. Classification of Mammals Above the Species
Level. Columbia University Press, New York.
Savage,
R.J.G. & Long, M.R. 1986. Mammal Evolution: an illustrated guide. Facts on File, New York.
Young, J. Z. 1977. La vida de los vertebrados. Editorial
Omega, Barcelona, 660 pp.
Paleontología en el Siglo XVIII y comienzo de la
anatomía comparada y el concepto de extinción.
Por Mariano Magnussen Saffer. Grupo Paleo. Museo
Municipal Punta Hermengo de Miramar. Fundación Argentavis.
marianomagnussen@yahoo.com.ar. Tomado de; Magnussen Saffer,
Mariano. 2016.
Paleontología en el Siglo XVIII y comienzo de la
anatomía
comparada y el concepto de extinción. Paleo Revista Argentina de
Paleontología. Boletín Paleontológico. Ańo XIV. 142: 18-19.
En su
obra Épocas de la Naturaleza, Georges Buffon se refiere a los
fósiles, en particular al descubrimiento de fósiles de especies
tropicales como los elefantes y rinocerontes en el norte de Europa, como
evidencia de la teoría de que la Tierra había sido mucho más cálida de
lo que actualmente era y se había ido enfriando gradualmente.
En
1796, Georges Cuvier presentó un documento sobre los elefantes vivientes
y los elefantes fósiles comparando los esqueletos de elefantes indios y
africanos con los fósiles de mamuts y de un animal que más tarde sería
denominado mastodonte utilizando anatomía comparada.
Cuvier
fue el primer gran promotor de la anatomía comparada y de la
paleontología. Ocupó diferentes puestos de importancia en la educación
nacional francesa en la época de Napoleón y tras la restauración de los
Borbones. Fue nombrado profesor de anatomía comparada del Museo Nacional
de Historia Natural de Francia, en París.
Estableció por
primera vez que los elefantes indios y africanos eran especies
diferentes, y que los mamuts se diferenciaban de ambos y por
tanto debían estar extintos. Incluso llegó a la conclusión de
que el mastodonte era otra especie extinta que también difería
de los elefantes indios o africanos, aún más que los mamuts.
Cuvier hizo
otra poderosa demostración del poder de la anatomía comparada en
la paleontología cuando presentó un segundo documento en 1796
acerca de un gran esqueleto fósil de Argentina, que nombró
Megatherium y se identificó como un perezoso gigante
comparando su cráneo al de las dos especies vivas de perezoso
arborícola.
El
innovador trabajo de Cuvier en paleontología y anatomía comparada
condujo a la aceptación generalizada de la extinción. También llevó
Cuvier a abogar por la teoría geológica del catastrofismo para explicar
la sucesión de organismos revelada por el registro fósil.
También
seńaló que, dado que ni los mamuts ni los rinocerontes lanudos
pertenecían a la misma especie que los elefantes y rinocerontes que
actualmente habitan en los trópicos, sus fósiles no podían ser
utilizados como evidencia de una tierra en enfriamiento gradual.
En una
aplicación inicial de la estratigrafía, William Smith, un
topógrafo e ingeniero minero, usó fósiles para ayudar a
correlacionar los estratos de suelo en diferentes ubicaciones.
Smith creó el primer mapa geológico de Inglaterra a finales de
los 1790 y principios del siglo
xix.
Estableció
el principio de sucesión faunística, la idea de que cada estrato de roca
sedimentaria contendría determinados tipos de fósiles, y que éstos se
sucederían entre sí de una manera predecible, incluso en formaciones
geológicas muy distantes.
Al mismo tiempo, Cuvier
y Alexandre Brongniart, instructor en la escuela de ingeniería minera de
París, utilizaron métodos similares en un influyente estudio de la
geología de la región circundante de París.
Bibliografía sugerida.
Bowler, Peter J. (2003).
Evolution:The History of an Idea. University of
California Press.
Bowler,
Peter J.; Iwan Rhys Morus (2005). Making Modern Science. The
University of Chicago Press.
Cadbury,
Deborah (2000), The Dinosaur Hunters: A True Story of Scientific
Rivalry and the Discovery of the Prehistoric World, Fourth Estate,
Desmond,
Adrian (1975). "The Discovery of Marine Transgressions and the
Explanation of Fossils in Antiquity". American Journal of Science,
Volume 275.
Palmer,
Douglas (2005) Earth Time: Exploring the Deep Past from Victorian
England to the Grand Canyon. Wiley, Chichester. I
Rudwick,
Martin J.S. (1985). The Great Devonian Controversy: The Shaping of
Scientific Knowledge among Gentlemanly Specialists. The University of
Chicago Press.
Zittel,
Karl Alfred von (1901). History of geology and palaentology to the
end of the Nineteenth Century. Charles Scribner's Sons,
London.
Wikipedia.
Que información deseas encontrar
en Grupo Paleo?
Nuestro sitio Web posee una amplia
y completa información sobre geología, paleontología,
biología y ciencias afines. Antes de realizar una
consulta por e-mail sobre algunos de estos temas,
utilice nuestro buscador interno. Para ello utilice
palabras "claves", y se desplegara una lista de
"coincidencias". En caso de no llegar a satisfacer sus
inquietudes, comuníquese a
grupopaleo@gmail.com