|
|
|
Fósiles del Oligoceno
de Argentina: |
|
|
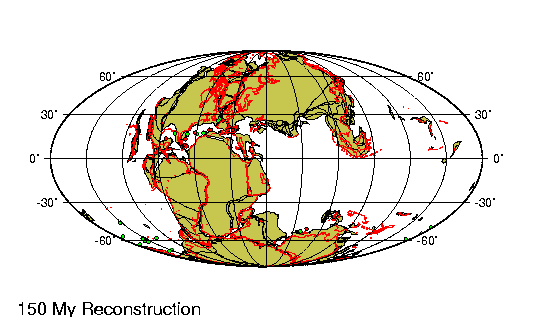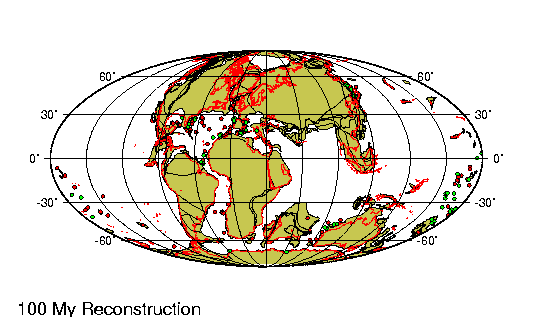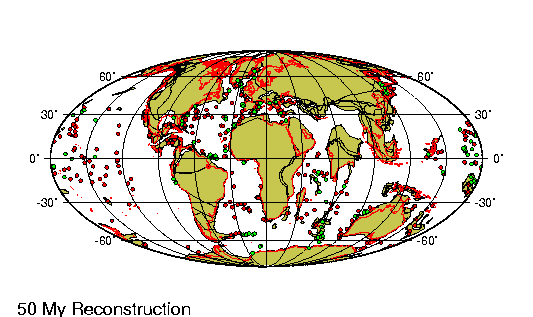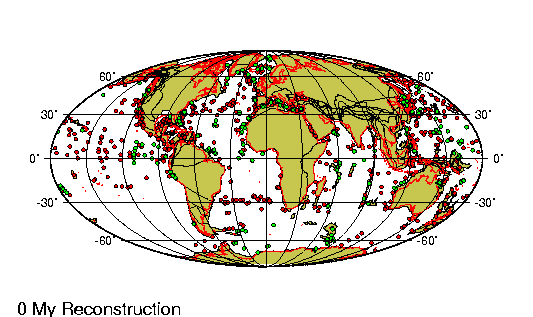
Se encuentra
representado en la República Argentina solo por dos edades y
comprenden de 35 a 24 millones de ańos antes del presente.

Sedimentos de Puerto Madryn.
|
 |
La edad mamífero
"Tinguiririquense", cuyos sedimentos fueron depositados en
un lapso intermedio del Eoceno y el Oligoceno, y comprenden
de una antigüedad de 36 a 31,5 millones de ańos. La segunda
edad es "Deseadense", en el cual desaparecieron una cantidad
considerable de mamíferos típicos de la región patagónica,
lo que pudo ser causado por el notable descenso del nivel
marítimo, calculados en unos 130 metros aproximadamente.
Para entonces el clima era templado, incluso en las tierras
más australes del continente. La presencia de numerosos
troncos silificados e improntas de vegetales ponen en
manifiesto la presencia de grandes bosques sub-tropicales.
Esta edad comprende entre los 29 y 24,5 millones de ańos.
Aparte de los afloramientos de patagonia, se observan otros
menos espectaculares en las Provincias de Mendoza y
Corrientes. A comienzos del Oligoceno las condiciones
climáticas se desarrollaron ostensiblemente. El clima era
templado y de vegetación estaba constituida por sabanas
arboladas y gramíneas. A mediados de esta época, los Andes
centrales se elevaron formando verdadero cordón montańoso
con gran actividad volcánica. Simultáneamente se produce un
brusco descenso en el nivel del mar, lo que debió exponer
gran parte de la plataforma continental. A raíz de esto
comienza a desarrollarse una leve deriva entre Tierra del |
|
Fuego y la Península Antártica que lleva la apertura del
Pasaje de Drake. A fines del Oligoceno el clima vuelve hacer
cálido, y las aguas oceánicas profundas y superficiales
vuelven a calentarse. Las condiciones en el continente eran
benignas y las floras estaban representadas por bosques,
sabanas arboladas y extensos pastizales. La plataforma
continental estaba cubierta por un mar relativamente somero
y de aguas templadas.
<<<
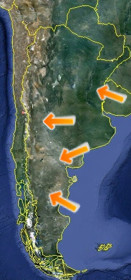
Principales sitios
fosilíferos de Argentina. |
|
Presbyornis pervetus. Wetmore,
1926. |
|
Presbyornis
Presbyornis
Presbyornis
Presbyornis
Presbyornis
Presbyornis
Presbyornis
Presbyornis
Presbyornis
Presbyornis
|
|


 |
|
|
|
Ave,
Presbyornithidae.
Es un género extinto de aves anseriformes. Está representada
por dos especies unánimemente aceptadas, el conocido
Presbyornis pervetus y el más desconocido
Presbyornis isoni. Justamente Presbyornis
pervetus tenía un tamańo similar al de un ganso,
pero con las patas más largas. Presbyornis isoni,
conocido a partir de unos pocos huesos, era más grande,
superando el tamańo del cisne. El Presbyornis es uno
de los anseriformes más primitivos. A causa de su cuello y
sus patas alargadas, al principio se pensaba que se trataba
de un flamenco, pero más adelante fue clasificado dentro de
los anseriformes cuando se descubrió la anatomía del cráneo
y del pico, muy similar a la de los patos. Más tarde, se
pensó que representaba una transición entre los anseriformes
y los charadriidae, pero se le considera parte de un grupo
extinto de anseriformes, los Presbyornithidae, relacionados
con los patos y los gansos. Según los hallazgos fósiles, se
cree que el Presbyornis vivía en colonias
alrededor de lagos poco profundos. Utilizaba su gran pico
para filtrar los alimentos del agua, que solían ser pequeńos
animales y plantas, como los patos actuales. |
Andrewsornis abbotti. Patterson,
1941.
|
 |
|
 |
|
Cráneo recreado a partir de los restos encontrados en la
Quebrada Fiera, inmediaciones de
Malargüe.
Museo de la ciudad de Mendoza. |
|
Aspecto de la
gigantesca ave hallada en el Oligoceno de la Provincia de
Mendoza. (*) |
Ave,
Phorusrhacidae.
Vivió en Oligoceno medio y
superior de Argentina. Esta ave de gran tamańo tenía 1,70 metros de
altura. Reconocido como un Phorusrhacidae, es un clado extinto de
aves cariamiformes conocidas también como “aves del terror”, porque
las especies más grandes eran depredadores en las diversas regiones
que habitaron. Eran aves carnívoras y no voladoras, siendo los
depredadores dominantes en América del Sur durante el Cenozoico,
entre 62 y 2,5 millones de ańos. Sus alas habían evolucionado para
utilizarlas como brazos para voltear una presa en movimiento. Eran
corredores rápidos. Sus parientes más cercanos hoy en día son las
chuńas de la familia Cariamidae. Restos de esta ave fueron exhumados
en el Oligoceno de Quebrada Fiera, inmediaciones de Malargüe,
Provincia de Mendoza. Su pico, grande y tosco le sirvió para pegar
cabezazos a su presa mientras la perseguía. Una vez reducido, con la
punta de su pico atravesaría los cráneos de su víctima, para luego
comenzar a comerla.
Physornis fortis.
Ameghino, 1895.
|
 |
|
 |
|
Imagen ilustrativa de Paraphysornis brasiliensis,
emparentado directamente con Physornis fortis del
Oligoceno de Argentina. Esqueleto montado en el MEF. |
|
Aspecto en vida
de Physornis fortis. (*) |
Ave,
Phorusrhacidae.
Fue una especie extinta de aves gigantes no
voladoras depredadores de la familia Phorusrhacidae o "aves del
terror", dentro de la subfamilia Brontornithinae, que vivían en
Sudamérica. Vivió entre los 28 a 23 millones de ańos, durante el
Oligoceno. Se ha encontrado exclusivamente en la región de Cabeza
Blanca, en Chubut. Su altura hasta la cabeza fue de aproximadamente
2 metros y el cráneo tenía una longitud de 60 centímetros. Se
alimentaban de mamíferos y probablemente de otras aves terrestres
parecidas al ńandú. Sólo competían con unos pocos depredadores como
cocodrilos y marsupiales similares al tigre diente de sable. Pesaba
unos 20 kilogramos, y vivía en lugares asociados a palmeras y
cuerpos de agua. Los fororracos vivieron después de los dinosaurios,
desde el Paleoceno, hace 50 millones de ańos, y hasta el
Pleistoceno, hace 1,8 millones de ańos. Su extinción, estaría
relacionada a cambios ambientales y climáticos, aunque
posteriormente fue reemplazado por una importante variedad de aves
del terror, que llegaron a medir unos 3 metros de altura durante el
Mioceno.
|
Anthropornis nordenskjoeldi.
Wiman, 1905. |
|
Anthropornis
Anthropornis
Anthropornis
Anthropornis
Anthropornis
Anthropornis
Anthropornis
Anthropornis
Anthropornis |
 |
 |
 |
|
Restos fósiles del pingüino
Anthropornis nordenskjoldi hallados en Tierra
del Fuego. (*) |
Humero de
Anthropornis sp. Museo de La Plata. |
Aspecto del primitivo pingüino
Anthropornis nordenskjoldi del Oligoceno del sur Patagonico.
(*) |
Ave.
Los pingüinos actuales constituyen un grupo muy
particular de aves no voladoras, adaptadas para la natación y buceo,
preferentemente en aguas frías y templadas. Su distribución
geográfica se restringe al hemisferio sur en varias regiones entre
la Antártida y las islas Galápagos. De manera semejante a su
distribución actual, los hallazgos de pingüinos fósiles también
están restringidos al hemisferio sur, y en particular el
conocimiento paleontológico de este grupo de aves está basado en
material fósil registrado en Nueva Zelanda, Australia, Sudáfrica,
Antártida y Argentina. Los pingüinos fósiles más antiguos,
aproximadamente entre 50 y 40 millones de ańos, se conocían para
Nueva Zelanda y Antártida. Algunos de ellos fueron verdaderos
gigantes y alcanzaron una talla entre 1,6 y 1,80 metros, como el
Anthropornis nordenskjoeldi, una especie fósil hallada en
la isla Marambio (Seymour), Antártida Argentina, por la célebre
expedición de Otto Nordenskjöld en 1903, en la cual también
participó el argentino José M. Sobral. En la actualidad la isla
Marambio, situada en el extremo norte de la península Antártica, se
encuentra separada del continente americano por el pasaje de Drake,
y aproximadamente a unos 1.200 km de distancia de Tierra del Fuego.
Sin embargo, hace unos 40 millones de ańos, el pasaje de Drake no
existía y la península Antártica estaba físicamente conectada al
extremo austral de América del Sur. El hallazgo en Tierra del Fuego
de pingüinos fósiles de edad semejante a la del material de la isla
Marambio en Antártida es consistente con esta antigua geografía y
apoya otras evidencias climáticas del pasado, por ejemplares el
marcado incremento en la abundancia de polen de Nothofagus
y las muestras isotópicas en conchillas marinas, que indican el
comienzo de una larga y continua tendencia de enfriamiento de las
aguas oceánicas australes hacia los 40 millones de ańos. A pesar de
la asociación popular de los pingüinos con las frías regiones
polares, existen especies de estas aves, que viven también más cerca
del ecuador. Los más primitivos, por tanto, debieron haberse
desarrollado en climas más cálidos, para adaptarse lentamente a sus
hábitats cuando éstos se helaron.
Parastrapotherium martiale. Ameghino 1901.
|
 |
|
 |
|
Rama
mandibular y pieza tipo de Parastrapotherium,
colectada por Ameghino y exhibida en el MACN. |
|
Reconstrucción del posible
aspecto (*) |
Mamífero
Astrapoterio.
Es uno de los elementos faunístico más
característicos de la Edad Mamífero Deseadense (Oligoceno tardío) de
Patagonia. Parastrapotherium reveló que este género
sólo presenta un premolar inferior más que Astrapotherium,
el cual es muy semejante a este último. Los astrapoterios,
constituyen un orden reducido, pero bien representado de herbívoros
de cuerpo bajo, que sobrevivieron desde fines del Paleoceno hasta
mediados del Mioceno. Algunos de ellos eran tan grandes como los
rinocerontes, con colmillos y una trompa pequeńa. Era herbívoro,
alimentándose principalmente de raíces, las cuales encontraba a
orillas de los paleopantanos donde pasaban la mayor parte de su
vida. Poseía un par de caninos bien desarrollados y una pequeńa
trompa. También se conoce a Parastrapotherium holmbergi
y Parastrapotherium ephebicum.
|
Thoatherium minusculum.
Ameghino, 1887. |
|
Thoatherium Thoatherium Thoatherium Thoatherium Thoatherium Thoatherium Thoatherium Thoatherium Thoatherium
|

Cráneo completo del extrańo Thoatherium
minusculum del
Oligoceno y Plioceno. Al lado, extremidad semejante a los
equinos actuales. (*) |
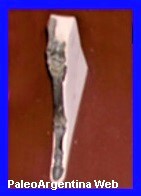 |

Reconstrucción del Thoatherium. (*) |
Mamífero
Litopterno.
Su nombre significa bestia feroz, y
fue descrito por Florentino Ameghino en 1887, a partir de restos
fósiles procedentes del Oligoceno y Mioceno de Patagonia. Es otro
ejemplo de evolución paralela, es decir, el desarrolló de animales
similares de antepasados diferentes, como es, la de los Litopternos
de América del Sur y los caballos verdaderos. Mientras que las
formas primitivas de equinos como Pliohippus se
convertían en caballos modernos en América del Norte, el
Thoatherium era un Liptoterna, un grupo de ungulados
sudamericanos nativos,
mucho mejor en varios aspectos. Como los caballos, el
Thoatherium tenía una sola pezuńa en cada pata, pero no hay
una sobrecańa que aun mostraba Hyracotherium. El
Thoatherium tenía las patas, el cuerpo, y la cabeza muy
parecida al de un caballo. Tenía una longitud de 70 centímetros,
siendo este, el de menor medida conocido entre los Litopternos.
Debido a sus largas patas, creemos que fue un excelente corredor.
Thoatherium probablemente consumía hojas y vegetación suave.
Cramauchenia normalis. Ameghino,
1902.
|
 |
|
 |
|
Cramauchenia insolita en el Field Museum of Natural History. |
|
Aspecto de Cramauchenia. Por John Conrad Hansen |
Mamífero
Litopterno.
Es
un género extinto de mamífero herbívoro, miembro del orden de los
litopternos, que vivió en América del Sur entre el Oligoceno
superior y el Mioceno inferior. Cramauchenia fue
nombrado por Florentino Ameghino. Su nombre no tiene traducción
literal; de hecho, es un anagrama del mejor conocido género
Macrauchenia. El género fue conocido inicialmente de la Formación
Sarmiento en la provincia de Chubut, en Argentina, siendo más tarde
descubierto en la Formación Chichinales en la provincia de Río Negro
y la Formación Cerro Bandera en Neuquén, también en Argentina, en
sedimentos asignados a la edad-mamífero sudamericana Colhuehuapense
(Mioceno inferior). En 1981 Soria hizo de C. insolita
un sinónimo menor de C. normalis. En 2010 se describió
un C. normalis encontrado en Cabeza Blanca (Chubut,
Argentina) en la Formación Sarmiento, en sedimentos asignados a la
edad-mamífero sudamericana Deseadense (Oligoceno superior).
Archaeohyrax patagonicus. Ameghino
1897.
|
 |
|
 |
|
Cráneo y mandíbula de
Archaeohyrax
patagonicus,
hallado en Chubut. Colección del Museo Argentino de Ciencias
Naturales de Buenos Aires. |
|
Posible
aspecto de Archaeohyrax
patagonicus (*) |
Mamífero
Notoungulado. Fue una especie de
mamífero notoungulado extinto, conocido en sedimentos desde el
Eoceno medio al Oligoceno de Argentina y Bolivia. Pertenecen a la
familia de los Archaeohyracidae, eran animales pequeńos poco
diversificados en un principio. Se les ha considerado en una
posición basal dentro del orden de los notoungulados y con todas las
características de los ungulados primitivos. Pasaron de tener el
aspecto y ocupar el nicho ecológico de los damanes. Se supone que
solían vivir en grupos y se comunicaban con sus congéneres mediante
diversos gritos. Su oído estaba muy desarrollado. Se alimentaban de
distintos vegetales dependiendo del tipo de especie.
|
|
Gualta cuyana. Cerdeńo, et, al.
2014. |
|
|
|
 |
 |
|
Maxilar in situ de
Gualta cuyana. Imagen del Conicet Mendoza. |
Aspecto de Gualta cuyana, según MarMag.2025. |
|
|
Mamífero
Notoungulado.
Fue un enorme mamífero perteneciente
al orden notoungulata. Algunos de sus fósiles fueron
encontrados en sedimentos terciarios de la Quebrada Fiera en
Malargüe, Mendoza. De este animal del Oligoceno tardío, se
han recuperado numerosos restos, entre ellos, el humero,
articulaciones, varias vértebras, cráneo y mandíbula, lo que
permite una reconstrucción de su aspecto en vida. Los
leontínidos eran animales con una estructura corporal
poderosa y es posible que algunos tuviesen cuernos como los
rinocerontes. Habitaron en Sudamérica, ocupando los nichos
ecológicos que en otros partes del mundo estaban ocupados
por los paquidermos y otros herbívoros de gran tamańo. Estas
bestias herbívoras eran cuadrúpedos. La familia se
identifica por poseer incisivos parecidos a los de los
cánidos. El resto de la dentadura suele ser braquiodonte. El
estudio comparativo permite diferencias morfológicas con
respecto a otros taxones del deseadense. Géneros
relacionados; Scarrittia, Leontinia, Ancylocoelus,
Anayatherium y Elmerriggsia. |
|
|
Leontinia gaudryi. Ameghino,
1895. |
|
Leontinia Leontinia Leontinia Leontinia Leontinia Leontinia Leontinia Leontinia Leontinia Leontinia Leontinia
|
 |
|
 |
|
Cráneo de Leontinia
gaudryi en el Amherst Museum of Natural
History. |
|
Aspecto de Leontinia
gaudryi. Tomado de deviantart.com. |
|
|
Mamífero
Notoungulado.
Es un género extinto de leontínido,
familia del orden de los
notoungulados que evolucionaron aisladamente
en Sudamérica. Florentino Ameghino fue quien, al describir el género
en 1895, le puso ese nombre en honor a su esposa parisina Leontina
Poirier. Sus fósiles se han hallado en la formación Deseado en
Argentina, y son de los más abundantes restos de cualquier animal
encontrados allí. El género data de finales del Oligoceno. En
Leontinia, los primeros dientes incisivos de la mandíbula superior
eran pequeńos dientes recortados, mientras los segundos eran más
alargados y formaban colmillos caniniformes. En la mandíbula
inferior el tercer incisivo se desarrolló como colmillo, no así el
segundo. Variaciones en la longitud de los incisivos han sido usados
para distinguir a las distintas especies del género unas de otras, y
podrían ser evidencia de dimorfismo sexual entre los individuos de
estas especies. De hecho, se ha sugerido que las distintas especies
asignadas sobre la base de las proporciones dentales podrían estar
realmente representando las variaciones de género entre los miembros
de una única especie. Los huesos nasales están muy elevados desde la
cavidad nasal y sugieren que este animal puede haber poseído una
especie de cuerno similar al del rinoceronte Diceratherium,
del Mioceno de América del Norte, Europa y Asia. |
|
Scarrittia canquelensis. Simpson, 1934. |
|
Scarrittia
canquelensis
Scarrittia
canquelensis Scarrittia canquelensis Scarrittia canquelensis
Scarrittia
canquelensis Scarrittia |
 |
 |
 |
|
Esqueleto Scarrittia
(*) |
Cráneo de
Scarrittia.
Foto Conicet |
Aspecto de
Scarrittia canquelensis
(*) |
|
Mamífero Notoungulado.
Es una especie extinta de mamíferos
notoungulados, el único miembro de los leontínidos que conocemos a
través de un esqueleto en buen estado de conservación. El nombre del
género, hace referencia a Scarrit Pocket, una cantera fosilífera
ubicada en la provincia de Chubut. Durante su vida, es probable que
se pareciera mucho a un rinoceronte de movimientos lentos y pies
planos. Scarrittia era un animal bastante pesado, de
cuerpo y cuello largos, patas robustas, pies con tres dedos
ungulados y un rabo muy corto. La tibia y el peroné estaban fundidos
parcialmente por la parte superior, por lo cual las patas no podían
girar hacia los lados. La cara era bastante corta y las mandíbulas
presentaban una dentadura completa, formada por 44 piezas con
coronas bajas y bastante poco especializadas. Scarrittia
canquelensis, Scarrittia barranquensis y Scarrittia
canquelensis, fueron encontradas en varias localidades de
Patagonia, mientras Scarrittia robusta, solo se
encontró en Paso del Cuello, en Uruguay. Este herbívoro de cuerpo
grande, y pesado tenía 2 metros de longitud. Su aspecto, tal vez,
era parecido al de un tapir, alimentándose de la floresta húmeda,
cerca de la costa de pantanos, ríos y lagos, en donde había
vegetación blanda, pastos, frutos y arbustiva.
|
Pachyrukhos mayonoi. Ameghino,
1885. |
 |
 |
 |
|
Esqueleto de Pachyrukhos
mayonoi. (*) |
Cráneo de Pachyrukhos
mayoni. (*) |
Recreación de Pachyrukhos.
(*) |
Mamífero Notoungulado.
Es una especie de
mamífero extinto, del orden de los notoungulados, es decir, los
extintos ungulados sudamericanos. Fue un pequeńo mamífero común en
la Patagonia a finales del Oligoceno y principios del Mioceno, es
decir, hace 25 millones de ańos atrás. Presenta morfología similar a
los roedores y media unos 30 centímetros de largo. Tenía una cola
muy corta y extremidades delanteras cortas a comparación de las
traseras. Su cráneo era ancho y tosco por la nuca, y se achicaba
hacia los nasales. Poseía mandíbulas fuertes, las cuales estaban
adaptadas para vegetales muy duros. Bulas timpánicas bien
desarrolladas, lo que podría seńalar orejas finas y largas. Cuencas
oculares algo grades seńalando hábitos crepusculares o nocturnos.
Pyrotherium romeroi. Ameghino, 1889.
|

|
 |

|
|
Cráneo de Pyrotherium
en el MEF. |
Mandíbula. Foto
Conicet. |
Reconstrucción
de Pyrotherium romeroi. (*) |
Mamífero
Pyrotheria.
Es una especie extinta de mamífero del
orden pyrotheria, propia de América del sur. Tenía unos 3,6 metros
de largo y pesaría 600 kilogramos. Su nombre significa "Bestia de
fuego" ya que los primeros restos hallados por Florentino Ameghino
en 1889, provenían de depósitos sedimentarios de origen volcánicos.
Algunos mamíferos fósiles de Sudamérica eran extrańos, pues,
evolucionaron aislados del resto de los continentes. Pyrotherium se
veía como un elefante primitivo en algunos aspectos, pero poseía
características propias muy llamativas. Las características nasales
del cráneo, de unos 75 centímetros, sugiere que desarrollo una
trompa, menor a la de los proboscideos. Tenía patas muy fuertes para
soportar el enorme peso de su cuerpo. Su cabeza estaba dotada de una
trompa y sus mandíbulas estaban acompańadas por seis colmillos, no
como los elefantes modernos o como algunas formas fósiles. Eran
cortos y en forma de cincel, los cuales eran utilizados para excavar
la tierra en busca de raíces y bulbos. Los esqueletos se conocen
imperfectamente, las especies más comunes halladas muestran un
cuerpo alargado y bajo. Algunas características recuerdan al grupo
de grandes herbívoros primitivos, los xenungulados del Paleoceno,
con los que se supone que están emparentados. Sin embargo, el
estudio completo del tarso de Pyrotherium no apoya
esta relación. No se observaron las características más derivadas
del mismo en otros mamíferos examinados, exceptuando al
Arsinoitherium del Paleógeno de África. Sus restos, fueron
también hallados en otros países de Sudamérica, como Brasil, Bolivia
y Perú.
Nesodon conspurcatus. Ameghino, 1887.
| |
|
|
|
 |
 |
 |
|
Vista inferior del cráneo
in-situ. (*) |
Cráneo
completo de Nesodon en el MEF. |
Aspecto del extinto
Nesodon. (*) |
Mamífero
Notoungulado. Es un género extinto
de mamíferos notoungulados de la familia Toxodontidae que vivieron
en el Oligoceno tardío al Mioceno medio. El aislamiento de América
del Sur, permitió la evolución separada de estos grupos de
mamíferos, muchos de los cuales se diversificaron en nichos
ecológicos que, en otras partes del mundo, ocuparon otros grupos. El
Nesodon se parecía a los rinocerontes y los hipopótamos. La
relación entre ellos se confirma tanto por la distribución de las
cúspides de los dientes como por los huesos del oído, que son
exclusivos de este orden de mamíferos. Nesodon se
tenía el cuerpo robusto. Alcanzaba un tamańo relativamente grande,
pesando más de 600 kilogramos, con 1.5 metros de altura a los
hombros y algo más de 3 metros de largo. El aspecto de estos
animales era impresionante. Su cuerpo era parecido al de un bisonte,
con la espalda arqueada y dotada de grupa. Las pezuńas eran pequeńas
y la cabeza de gran tamańo, casi desproporcionada con el resto del
cuerpo. Se alimentaba de vegetales en zonas áridas. Es una de las
especies más características del Oligoceno de Patagonia Argentina.
|
Anatrachytherus soriai. Rugero, M. 2004. |
|
Anatrachytherus Anatrachytherus Anatrachytherus Anatrachytherus Anatrachytherus Anatrachytherus Anatrachytherus
|
|
Ilustraciones
mandibulares y reconstrucción de
Anatrachytherus soriai, del
Oligoceno de Patagonia.
(*) |
Mamífero
Notoungulado.
Los Trachytheriinae son mamíferos
notoungulados mesotéridos de tamańo mediano, similar al de un
cordero, con un peso estimado de aproximadamente 60 kilogramos. Se
consideran herbívoros pastadores por la gran altura de sus
molariformes y la espesa capa de cemento que cubre los mismos. La
distribución cronológica de los Trachytheriinae es Edad
divisaderense-Edad deseadense. Ameghino en 1897, describió e ilustró
una mandíbula con la dentadura completa, proveniente de Cabeza
Blanca, Provincia de Chubut, descubierta por Carlos Ameghino en
1896, y la asignó a Trachytherus spegazzinianus =
Anatrachytherus soriai, la única especie de
Trachytheriinae reconocida para el Deseadense de Patagonia. Los
sedimentos portadores han sido referidos a la Formación Sarmiento.
Representantes de la subfamilia Trachytheriinae son conocidos en
Argentina y en Bolivia. Sin embargo, hay registros extrapatagónicos
en las provincias de Mendoza y Corrientes como el traquiterino de la
Formación Divisadero Largo en Mendoza, conocida como
Trachytherus mendocensi.
Trachytherus spegazzinianus. Ameghino, 1889.
|
 |
 |
|
|
Cráneo de un
Trachytherus s.p (ilustrativo) del Museo de Historia
Natural de Lima. Oligoceno de Perú. |
El técnico
Mariano Magnussen Saffer, junto a restos de
Trachytherus spegazzinianus,
recreados a partir de un cráneo del oligoceno de Patagonia.
Museo de Cs Nat de Miramar. |
Aspecto de
Trachytherus s.p. (*)
|
Mamífero
Notoungulado.
Se trata de otra especie de mamífero
ungulado nativo de Sudamérica, es decir herbívoros con pezuńas. Este
notoungulado mesoterio según todos los indicios era un animal con
pezuńas muy diferente. Tenía dientes de crecimiento permanente
parecidos a los que presentan los roedores. Adicionalmente su
distribución parece circunscribirse solamente al Oligoceno tardío y
su distribución estratigráfica se restringe a la parte superior del
miembro Puesto Almendra de la formación Sarmiento, en la cuenca del
golfo de San Jorge, provincia del Chubut, y a los estratos de la
cuenca de Salla-Luribay, Bolivia. En Patagonia, Pyrotherium
romeroi tiene la misma distribución estratigráfica que la de
Trachytherus spegazzinianus. Se ha recomendado también
a esta especie para caracterizar la fauna del Oligoceno asignada a
la edad deseadense en la Patagonia y Bolivia, al contar con un
completo registro estratigráfico. Era un animal de mediado tamańo,
semejante a una oveja. Usaban sonidos para comunicarse entre ellos,
posiblemente para emparejarse y advertir del peligro. Eran
primariamente herbívoros y se desplazaban según su hábitat, en busca
de hojas, raíces, flores, cortezas, frutos, etc, ya que, la
Patagonia no era un gran desierto como en la actualidad, sino era
más bien, una selva subtropical. Adicionalmente, también comían
insectos y pequeńos animales.
|
Rhynchippus equinus. Ameghino,
1897. |
|
Rhynchippus equinus
Rhynchippus equinus
Rhynchippus equinus
Rhynchippus equinus
Rhynchippus equinus |
|

 |
|
|
|
Mamífero
Notoungulado.
Es un género extinto de mamíferos
notoungulados de la familia Notohippidae, que vivió a principios del
periodo del Oligoceno en el continente sudamericano. Sus fósiles se
han hallado en Argentina y Chile. Representa un ejemplo clásico de
la convergencia de los ungulados sudamericanos con otros grupos no
relacionados del resto del mundo, en este caso, con el caballo. Era
muy similar al antepasado de los caballos Hyracotherium
del período Eoceno. Era un pequeńo herbívoro del tamańo de un zorro,
con un metro de longitud, parecido a los caballos actuales que el
propio Hyracotherium. Tenía tres dedos en las patas
delanteras, en vez de los cuatro del hiracoterio, y tres en las
traseras, protegidos por pezuńas, siendo el central más largo. Su
esqueleto, con el cuerpo grueso y los dedos en forma de garra, no se
parecía demasiado al del caballo, pero los dientes eran similares a
los de un animal que se alimenta de hierba, como el caballo o el
rinoceronte. Los caninos no formaban colmillos, como en la mayoría
de los demás toxodontos, sino que tenían el mismo tamańo y la misma
forma que los incisivos. Eran altos e ideales para cortar plantas.
Las muelas eran grandes y llevaban esmalte enrollado para moler la
vegetación dura y, además, estaban recubiertas de cemento. A pesar
de que alguna vez se creyó que eran los antepasados de los caballos
auténticos, las similitudes, que residen fundamentalmente en la
forma del cráneo y en los incisivos para cortar plantas, son
consecuencia de la evolución convergente, es decir, formaron
características morfológicas similares debido que evolucionaron en
ambientes parecidos. En lo esencial, presenta todas las
características de los notoungulados.
|
|
|
Mendozahippus fierensis. Cerdeńo y
Vera, 2010. |
|
|
|

 |
|
Cráneo de
Mendozahippus, tomado de morphobank. |
Posible aspecto de
Mendozahippus (*)
|
|
|
|
|
Mamífero
Notoungulado.
Es una especie de mamífero
notoungulado extinto, que existía en la región cuyana, en la
provincia de Mendoza, durante el Oligoceno tardío. Los
fósiles se conocen desde el sitio de la Quebrada Fiera del
Agua de la Formación Piedra e incluye una completa cráneo y
dos asociados metatarsianos, dos maxilares fragmentos y
cinco dientes superiores aisladas. Los notohípidos (Notohippidae,
griego "caballos meridionales") son una familia extinta de
mamíferos placentarios del orden Notoungulata, perteneciente
al superorden Meridiungulata, que habitaron en Sudamérica.
Fueron unos cuadrúpedos herbívoros con caracteres que
indican un cambio de hábitat del bosque a las praderas. Las
patas fueron alargándose y la dentadura muestra que se
alimentaban cada vez con mayor frecuencia de pasto que de
follaje. Los fósiles de los notohípidos están datados desde
el Eoceno hasta el Oligoceno tardío, es decir de hace unos
25 millones de ańos antes del presente. En esta época, la
cordillera de los Andes no se había elevado y el clima era
diferente. Entre los restos encontrados de estos mamíferos
herbívoros, se incluyen, entre otros, los hegetoterios tan
pequeńos como conejos, los notohípidos, con aspecto de un
caballito del tamańo de un perro grande, los leontínidos,
que doblan en tamańo a los anteriores, piroterios, con el
aspecto y la talla de un elefante mediano y homalodoterios,
quizá los más parecidos a los conocidos rinocerontes. A
ellos les acompańaban depredadores como marsupiales
carnívoros y grandes aves corredoras, los fororracos.
Santiagorothia chiliensis. Hitz et al. 2000.
|
 |
|
 |
|
Cráneo de
Santiagorothia chiliensis
(*). |
|
Aspecto en vida de
Santiagorothia chiliensis.
Por
Darin Croft and Velizar Simeonovski |
Mamífero
Notoungulado. Fue una especie de
mamífero notoungulado, interatheríido, herbívoro, que vivió
hace entre 40 y 28 millones de ańos, en el Oligoceno
Temprano. Sus restos fosiles fueron hallados en la provincia
de Río Negro, y descrito por Hitz. En el orden Notoungulata,
los interatéridos fueron animales en su mayoría de pequeńo y
mediano tamańo que ocuparon el hábitat de animales actuales
como las liebres o las vizcachas. La mayoría de los
interaterios eran mamíferos bastante pequeńos, parecidos a
los roedores. Fueron un grupo longevo, con representantes
fósiles desde fines del Paleoceno hasta fines del Mioceno.
La familia Interatheriidae es uno de los grupos de mamíferos
mejor representados en la fauna de la Formación Santa Cruz.
Sus cráneos también recuerdan vagamente a los roedores, otro
ejemplo de convergencia adaptativa, donde sus molares
demuestran estar adaptados para una dieta herbívora. Pudo
ver sido un gran corredor. Otro género bien conocido es
Interatherium, del cual el resto fósil más abundante
pertenece a Interatherium robustum. |
|
|
Cladosictis lustratus. Ameghino,
1887. |
|
Cladosictis Cladosictis Cladosictis Cladosictis Cladosictis Cladosictis Cladosictis Cladosictis Cladosictis Cladosictis
|
 |
|
 |
|
Cráneo del primitivo depredador marsupial
sudamericano Cladosictis lustratus, hallado en Santa Cruz. (*) |
|
Cladosictis lustratus,
un astuto y pequeńo marsupial del Oligoceno de la Patagonia
Argentina. (*) |
Mamífero
Marsupial. Fue un depredador
marsupial con aspecto de coipo (falsa nutria) por su forma y pequeńa
dimensión, con un cuerpo largo y extremidades relativamente cortas,
con un tamańo de la cabeza a la cola de 80 centímetros. La hembra
daba a luz a pequeńos embriones, los cuales terminaban su desarrollo
en una bolsa materna externa. Era de hábitos carnívoros,
alimentándose de peces, ranas, reptiles y mamíferos pequeńos. Los
dientes comprendían de incisivos en la parte anterior, para sujetar
las presas. Los caninos puntiagudos para matar, molares y premolares
carniceros. La fórmula dentaria de este extrańo marsupial hace
acordar a las de los mamíferos placentarios.
Proborhyaena gigantea. Ameghino, 1897.
Mamífero
Marsupial.
Fue un marsupial de hábitos
carnívoros, del orden Sparassodonta del tamańo de un lobo, de unos 2
metros de longitud. Era parecido a un oso, con las patas planas.
Como muchos metaterios, probablemente las hembras poseían una bolsa
para llevar a las crías en desarrollo. Los borhiénidos tenían
fuertes y potentes mandíbulas, capaces de triturar huesos. Eran
depredadores muy importantes durante el Oligoceno de la Patagonia
Argentina, los cuales se alimentaban de mamíferos notoungulados y
armadillos. Las patas cortas seńalan que no era un animal corredor,
pero seguro que asechaba a sus víctimas en los antiguos bosques que
existían hace 30 millones de ańos en los actuales desiertos de la
provincia de Chubut. Originalmente, la familia Borhyaenidae fue uno
de los grupos más extensos entre los esparasodontes, incluyendo a
todas las especies que no fueron incluidas originalmente en
Thylacosmilidae (marsupiales dientes de sable). Sin embargo, en ańos
recientes, otros grupos de esparasodontes han sido elevados al rango
de familia y esto junto al descubrimiento de que los borhiénidos
están más cercanamente relacionados con los proborhiénidos y los
tilacosmílidos que a otros esparasodontes, la familia ha sido
reducida a seis especies en tres géneros.
Palaeothentes pressiforatus. Ameghino 1887.
|
 |
|
 |
|
Cráneo y
mandíbula de Palaeothentes lemoinei, del Mioceno de Santa Cruz.
(*). |
|
Aspecto del
pequeńo Palaeothentes. Por MarMag.2025 |
Mamífero
Marsupial. Fue un mamífero
marsupial con características similares a las musarańas, endémicas
de América del sur, que incluye a géneros vivientes como
Caenolestes, Lestoros y Rhyncholestes,
caracterizados por su pequeńo tamańo y su dieta insectívora -
frutícola. Conocidos desde Eoceno temprano, pero con más presencia
durante el Oligoceno. Este pequeńo animal, no más grande que un
ratón, tenía una especialización particular en los dientes, similar
a los roedores. El Palaeothentes pertenece a la
superfamilia de Caenolestidae, mamíferos arcaicos marsupiales que
sobrevivieron con pocas especies hasta la actualidad. Entre las
especies más conocidas del género, podemos citar a
Palaeothentes pascuali, Probablemente Palaeothentes era un
excelente escalador.
Kramadolops maximus. Goin et al. 2010.
|
 |
|
 |
|
Fósil de Kramadolops maximus.
(*). |
|
Aspecto de Kramadolops maximus.
Por MarMag.2025 |
Mamífero
Marsupial. Es una especie extinta de mamífero marsupial
polidolópido. Eran pequeńos animales herbívoros, de dieta variada,
como frugívoro, insectívoro y folívoro. De estos marsupiales se
conocen principalmente restos de dientes, mandíbulas y maxilares. Se
estima que el tamańo pudo variar de acuerdo a la especie de cada
integrante de la familia, entre los 70 gramos y los 3,5 kilogramos.
Los polidolópidos se conocen del sur de América del Sur (en Chile y
la Patagonia Argentina) y de la Península Antártica. Los
polidolópidos antárticos fueron registrados en la Isla Marambio en
niveles de una edad aproximada de 45 Millones de ańos. Si bien hoy
parece impensable que un marsupial (y cualquier mamífero terrestre)
sobreviviera en este continente, en la época en la que vivieron los
polidolópidos, había grandes bosques similares a los bosques andino
patagónicos, y aun compatibles con climas más cálidos en tiempos más
antiguos. Más aun, durante gran parte del Mesozoico, América del
Sur, Antártida y Oceanía estuvieron unidos, y es por eso que muchos
grupos de plantas y animales hoy están compartidos en estos
continentes en la actualidad (o en el registro fósil). En
particular, hacia fines del Cretácico y comienzos del Paleógeno
(hace unos 66 Millones de ańos) el sur de América del Sur y la
Antártida estuvieron unidos por lo que los polidolópidos pudieron
haber evolucionado en esta gran masa continental sin tener mayores
barreras que las aislaran. Por eso hoy los registros de la Antártida
nos muestran restos de estos grupos (junto con otros grupos de
mamíferos). Incluso es posible que este grupo de marsupiales esté
más relacionado con los marsupiales australianos que con las
zarigüeyas sudamericanas actuales. Los polidolópidos desaparecieron
del registro a comienzo del Oligoceno, hace unos 30 millones de
ańos, probablemente debido al deterioro del clima, con una
disminución tanto de la temperatura como la humedad, particularmente
en la región patagónica donde estos restos fósiles fueron
registrados.
Los saltadores de Islas,
balseros y el intercambio faunístico con África.
 |
|
En el lago Chippewa,
en Wisconsin en, EE.UU.,, alberga una isla
flotante gigante que a veces se mueve y bloquea un
puente extremadamente importante y necesita ser
movida por barqueros locales.La masa flotante posee
árboles de gran tamańo y animales. |
|
A fines del Eoceno y
principios del Oligoceno, hubo un breve ingreso y variación
del contenido biológico conocido en la aislada Sudamérica,
la cual, funciono como un continente aislado luego de la
extinción de los dinosaurios, en el Cretácico, desarrollando
una fauna muy particular, que se vio perturbada en algunas
remotas ocasiones. De aquí surgen los primeros saltadores de
islas o balseros de origen africano.
Sobrevivir
a un viaje intercontinental sobre una masa de vegetación
parece una posibilidad entre un millón. Por ello, los
científicos han intentado determinar si hubo un único viaje
en balsa con roedores y monos juntos, sobre un lecho de
plantas enredadas, sedimentos, entre otros, que incluso hoy
en día, suelen darse estos fenómenos naturales. |
Los paleontólogos
aún están tratando de averiguar cuándo se produjeron exactamente
estas travesías. El nuevo estudio sugiere que las travesías tuvieron
lugar hace unos 40.5 millones de ańos, cuando América del Sur y
África estaban separadas por solo 965 kilómetros, mucho más cerca
que el golfo actual, de casi 3000 kilómetros. El nivel del mar
descendió, cerrando la distancia oceánica de una forma diferente.
Una importante erosión en entornos cercanos a la costa podría haber
provocado el desprendimiento de grandes balsas, repletas de animales
y plantas.
Los primeros
intercambios faunísticos que vivió Sudamérica acontecieron a finales
del periodo Eoceno hace aproximadamente 31,5 millones de ańos, con
la llegada de los ancestros de los roedores cávidos. La llegada de
estos primeros inmigrantes de origen africano marcó el fin para
muchos pequeńos marsupiales sudamericanos, y sentó las bases para la
evolución de los actuales capibaras, chinchillas, vizcachas y
ursones, como coendúes y puercoespines.
Poco después, hace
aproximadamente 25 millones de ańos, llegó una nueva oleada de
inmigrantes; esta vez los principales inmigrantes fueron los
ancestros de los monos americanos, los platirrinos. Estos pequeńos
primates llegaron a un continente con pocos animales arborícolas
capaces de hacerles competencia, lo que derivó en un fructífero
proceso de diversificación.
Estas migraciones
fueron posibles gracias a la relativamente corta distancia existente
entonces entre Sudamérica y la costa occidental de África, a la
existencia del archipiélago de San Pedro y San Pablo que, por su
ubicación, hicieron posible el salto entre continentes, y a la
existencia de fuertes corrientes oceánicas en dirección oeste.
En el Oligoceno,
llegaron nuevos inmigrantes. Los más destacados de esta oleada
fueron las tortugas terrestres que, en un principio, se creyó habían
venido de Norteamérica, pero recientes estudios de ADN han permitido
demostrar que las tortugas terrestres sudamericanas y las tortugas
gigantes de las islas Galápagos tienen un parentesco más cercano con
las tortugas del género Kinixys, oriundo del centro de África que
con las tortugas terrestres y dulceacuícolas de Norteamérica.
|
Tremacebus harringtoni. Rusconi,
1933. |
| Tremacebus Tremacebus Tremacebus Tremacebus Tremacebus Tremacebus Tremacebus Tremacebus Tremacebus Tremacebus
|
|

|

|

|
|
Cráneo de Tremacebus, un
primate primitivo procedente del Oligoceno. (*) |
Cráneo de Tremacebus harringtoni. (*)
|
Aspecto posible del primate
de la desaparecida selva patagonica. (*) |
Mamífero
Primate.
Fue un primitivo mamífero primate
(monos) aloctono, cuyo ingreso al continente fue hace 38 millones de
ańos según la evidencia fósil reunida hasta el momento. A diferencia
de los monos del Viejo Mundo, los monos del Nuevo Mundo no parecen
haberse dado a la vida en las praderas y siguieron siendo animales
de selva. Una gran diferencia entre ambos, es que los primeros no
poseen cola, mientras que los sudamericanos poseen unas vértebras
caudales extensas y adaptadas como un quinto miembro para poder
trepar. Tremacebus harringtoni, es un representante
del periodo Oligoceno tardío, es decir, de unos 24 millones de ańos
antes del presente, y se parecía a un mono búho moderno. Otros
pequeńos simios del territorio argentino son Homunculus
patagonicus y Killikaike blakei. Los primates
más primitivos por ahora en América del sur es Branisella, del
Oligoceno temprano, de unos 34 millones de ańos procedente de
Bolivia. Mientras que dos parientes del actual mono arańa, el
Protopithecus y el Caipora, vivieron en Brasil durante el
Pleistoceno. Tremacebus era omnívoro y tenía una
variada dieta, lo que lo convertía en un animal exitoso para
resistir cualquier cambio. Este género fue descrito originalmente
por el paleontólogo Hershkovitz en el ańo 1974. La única especie en
que está basado, fue descrita originalmente por el paleontólogo
Carlos Rusconi en el ańo 1933. Se incluye en la familia de los
aótidos (Aotidae), exclusiva del Neotrópico, que comprende un solo
género viviente, con un total de 11 especies, siendo los únicos
monos nocturnos. Carlos Rusconi recibió la muestra en el ańo 1932
enviada por Harrington Thomas, quien lo recogiera junto con otros
restos fósiles de mamíferos de la edad Colhuehuapense, encontrados a
unos 12 kilómetros al suroeste del cerro Sacanana, en el
centro-norte de la provincia del Chubut, en el centro de la
Patagonia argentina.
Dolichocebus gaimanensis. Bordas 1942.
|
 |
|
 |
|
Cráneo de Dolichocebus gaimanensis, de la colección del MACN.
Tomado del Twitter MACN.VP |
|
Posible aspecto de Dolichocebus gaimanensis. (*) |
Mamífero
Primate.
Es un primate platirrino que pertenece
al Oligoceno tardío y Mioceno temprano. Vivió en la Patagonia
argentina, cuando esta tenía una exuberante selva, a diferencia de
lo que vemos en la actualidad. Pesaba de 2 a 3 kilogramos, como el
actual género Cebus. Fósiles muy completos proceden del acantilado
sur del Valle Bajo del río Chubut, cerca de Gaiman y en Sacanana.
Dolichocebus se conoce por el tipo de cráneo encerrado en una
concreción, numerosos dientes aislados, partes de dos mandíbulas y
un astrágalo. Seguramente tenía una cola prensil que enrollan
alrededor de las ramas para ayudarse en el movimiento en los
árboles. Debió, adaptándose a varios tipos de bosques y consumiendo
muchos tipos de comida, que incluian frutas, diferentes vegetales,
invertebrados y pequeńos vertebrados. Los platirrinos o monos del
Nuevo Mundo se diferencian de los del Viejo Mundo, entre otras
características, por las fosas nasales, que están más separadas y
dirigidas hacia adelante en lugar de hacia abajo. Debido a lo pobre
de su registro fósil, las relaciones de parentesco de los monos
sudamericanos con otros grupos de primates y entre sí son las menos
conocidas dentro del orden de los primates. La mayor parte de los
platirrinos fósiles son conocidos solamente por dientes y mandíbulas
aisladas. De unos pocos se conocen cráneos, los que generalmente
carecen de dientes, están deformados o mal preservados.
Octodontotherium grandae. Ameghino, 1894.
|
 |
 |
|
Fragmento mandibular
atribuido a
Octodontotherium s.p del Oligoceno de
Chubut. Museo de La Plata. |
Posible aspecto de
Octodontotherium.
(*) |
Mamífero
Xenarthro.
Es un género extinto de perezoso
terrestre de la familia mylodontidae, endémica de América del Sur
durante el Oligoceno-Mioceno, poco común de hallar para estas
edades. Vivió entre 29 a 21 millones de ańos antes del presente,
existente por aproximadamente 8 millones de ańos. Era uno de los más
grandes edentados para su época, con una talla similar al del genero
Lestodon que vivió durante el Pleistoceno. Provisto de grandes y
gruesos huesos por lo poco que sabemos de esta especie. Su
alimentación era herbívora. El material asignado a Octodontotherium,
colectado en distintas campańas por E. Riggs a principios del siglo
XX y depositados en el Museos de Historia Natural de Chicago, como
así también, en París, La Plata y Buenos Aires, permitió el estudio
de numerosos elementos, como la tibia, el astrágalo, mandíbulas y un
cráneo bien conservado. La distribución de fósiles es exclusiva de
las provincias de Chubut y Santa Cruz, Argentina. Otra especie
relacionada; Orophodon hapaloides Ameghino, 1894.
Deseadognathus riggsi. Carlini y Scillato-Yané,
2004.
|
 |
|
 |
|
Ramas mandibulares de
Deseadognathus riggsi
del Oligoceno de la Provincia de Santa Cruz.
|
|
Posible
aspecto del
Megalonychidae
Deseadognathus (*). |
Mamífero
Xenarthro.
Es un género extinto de perezoso
terrestre Megalonychidae del Oligoceno de Patagonia. Pudo haberse
parecido a un típico perezoso terrestre megaloníquido, aunque era
mucho menor, con una cola alargada y un torso ancho, así como
miembros ágiles y largas garras. Los dientes caniniformes de
Deseadognathus riggsi eran grandes y triangulares, y su
cráneo era profundo y tenía una gran cresta sagital la cual en
conjunto con la mandíbula ancha probablemente permitía un fuerte
esfuerzo realizado por los músculos masticatorios, como otros
representantes de este grupo. Seguramente fue un animal solitario,
nocturno y arborícola, que habitaba en la selva secundaria y
desarrollada, en los bosques extintos de la Patagonia. El nombre
perezoso significa lento, pero los movimientos lentos de este animal
son en realidad una adaptación para sobrevivir con una dieta de baja
energía basada en hojas. Estos perezosos tienen la mitad de la
velocidad del metabolismo de un mamífero del mismo tamańo. Los
perezosos actuales, tienen una vista y oído muy pobre por lo que la
mayoría del tiempo se basan en tocar y oler para conseguir alimento,
por lo cual, es posible que sus representantes en estado fósil,
tuvieron características y compartimentos similares. Sus restos
proceden de la localidad de Puerto Deseado, provincia de Santa Cruz.
Pseudhapalops observationis. Ameghino, 1891.
|

|
 |

|
|
Esqueleto casi completo de Pseudhapalops
s.p, exhibido en el Museo de La Plata.
Oligoceno-Mioceno temprano de Santa Cruz. |
Copia de cráneo y mandíbula de Pseudhapalops
s.p, en el Museo de Ciencias
Naturales de Miramar. Oligoceno-Mioceno temprano de Santa
Cruz. |
Aspecto en vida
de Pseudhapalops
s.p, cuando la Patagonia era una selva tropical. (*) |
Mamífero
Xenarthro.
A finales del Oligoceno, hace 24
millones de ańos, los Xenarthros estaban bien representados y
adaptados en América del Sur, por lo cual, en el Mioceno se
encontraba geográficamente bien distribuido por la actual Patagonia
Argentina. Pseudhapalops era de tamańo más bien chico,
media hasta un metro de largo, a diferencia de sus gigantescos
parientes del Pleistoceno que superaban los cuatro metros de altura.
Tenía garras muy desarrolladas y curvas (una característica común
entre los mamíferos edentados) seguramente debió desplazarse sobre
sus nudillos anteriores. Es muy probable que pasara la mayor parte
de su tiempo en las copas de los árboles, sujetándose de sus garras
y alimentándose de frutos suculentos en los tupidos bosques sub
tropicales del Oligoceno patagónico. Como todos los edentados,
Pseudhapalops tenía pocos dientes y sin esmalte. Sus
mandíbulas solo tenían cuatro pares de dientes. Si bien hoy en día
la Patagonia Argentina es un gran desierto, durante el Oligoceno y
Mioceno estaba constituida por grandes y extensas selvas,
evidenciado por la diversidad faunistica y botánica recuperada.
Otras especies relacionadas; Eucholoeops ingens y
Pelecyodon s.p.
Peltephilus pumilus.
Ameghino, 1904.
|
 |
 |
|
Cráneo y mandíbula
de Peltephilus. MACN. |
Reconstrucción en vivo . (*). |
Mamífero
Xenarthros.
Fue una especie de mamífero xenarthro,
emparentado con los actuales armadillos (tatus, peludos y mulitas).
Tenía una coraza formada por placas óseas, las cuales habían
evolucionado de placas dérmicas, formando un caparazón recubierto de
córnea y con bandas móviles, lo que le brindaba movimientos rápidos.
Peltephilus, poseía en el hocico un par de cuernos
largos, originado de osteodermos modificados, protegiendo los ojos,
que en vida se encontraban recubiertos por piel y tejido corneo, lo
que facilitaba su utilización en momentos de defenderse de sus
depredadores. Algunos científicos sostienen que este armadillo,
tenía unos 60 centímetros de largo, y que, tal vez poseía otro par
de pequeńos cuernos córneos en el hocico. A diferencia de otros
edentados, Peltephilus, tenía unos dientes gruesos y
largos, muy parecidos a caninos, lo que podría indicar que era un
animal carnívoro o al menos, carrońero, conducta que se repitió en
otros armadillos más tardíos, aunque hay investigadores que
sostienen una dieta exclusivamente herbívora. Curiosamente, el
desarrollo de estos cuernos óseos, pareciera ser una convergencia
adaptativa que comparte con el roedor norteamericano
Ceratogaulus, siendo ambos, los únicos mamíferos fosoriales
que poseen cuernos. Seguramente, construyo sus propias madrigueras
para refugiarse o cuidar de sus crías.
Clypeotherium
magnum. ScillatoYané, 1977.
|
 |
|
 |
|
Coraza de
Clypeotherium magnum.
del Oligoceno de Chubut,
Argentina. Museo de La Plata. |
|
Posible aspecto de Clypeotherium magnum.
|
Mamífero
Xenarthros.
Se trata de un mamífero placentario
extinto. Este enorme armadillo es comparable con los que vivieron
durante el Pleistoceno, perteneciente a la familia extinta
Glyptodontidae, del orden Cingulata. Originarios de Sudamérica, se
diferenciaron en el Eoceno Superior. Tenían un cráneo y un caparazón
formado por muchas piezas pequeńas fusionadas y, en ocasiones, una
cola cubierta. Muestran una notable convergencia con las tortugas y
algunos dinosaurios (anquilosauros). Tenían una adaptación a la
dieta herbívora y conversión de sus bandas acorazadas del dorso en
una suerte de único caparazón defensivo. Perteneció a la subfamilia
de los Glyptatelinae, gliptodontes primitivos conocidos en el
Oligoceno de Argentina, que aparentemente pudieron sobrevivir de
manera relictual en el norte de Sudamérica en el Neógeno. Su coraza
tiene un grosor de 4 centímetros, y por el margen de esta, estaba
constituidas por placas puntiagudas. Su caparazón dorsal completo,
llegaba a los 3 metros de largo.
Palaeopeltis
inornatus. Ameghino 1894.
|
 |
|
 |
|
Placas u osteodermos de
Palaeopeltis inornatus (*). |
|
Posible
aspecto de
Palaeopeltis inornatus (*). |
Mamífero
Xenarthros.
Es una especie de mamífero extinto de
gliptodontes, grandes mamíferos xenartros emparentados con los
modernos armadillos con los que se incluyen en el orden Cingulata,
caracterizados por la presencia de corazas óseas a lo largo de su
cuerpo, incluyendo cabeza y cola, que vivieron en Sudamérica durante
el aislamiento geográfico. Los gliptodontes sin embargo, formaban
una familia diferenciada, ya que carecían de varias bandas móviles
como sus parientes actuales, siendo éstas fusionadas en un caparazón
más o menos rígido de pocas o ninguna banda en el dorso, además de
presentar adaptaciones para la dieta herbívora como el acortamiento
del cráneo, con la aparición de espinas en el hueso malar para
ayudar a sostener un músculo masetero de mayor tamańo y el
desarrollo de molares altos especializados en la masticación de
materia vegetal. Esta nueva clasificación mostró que la subfamilia
Glyptodontinae probablemente no evolucionó a partir de los
gliptodontes de las subfamilias Propalaehoplophorinae o
Hoplophorinae en el sur del continente durante el Plioceno, sino que
más bien apareció en el norte de Sudamérica a mediados del Mioceno y
luego avanzó hacia al sur, aprovechando los cambios climáticos que
favorecieron los ambientes de sabanas y que serían característicos
poco después, cuando se produjo el Gran Intercambio Biótico
Americano. Los Palaeopeltidae fueron por primera vez descritos por
Ameghino en 1894, a través de osteodermos provenientes de sedimentos
donde también se hallaron los restos de Pyrotherium, los cuales
corresponden a la “Edad Mamífero Deseadense” en la Patagonia
Argentina. También se encontraron restos de Palaeopeltis inornatus
para la Formación Fray Bentos, Oligoceno tardío de Uruguay.
Chubutolithes
gaimanensis. Bown & Ratcliffe 1988.
|
 |
|
|
 |
|
Nidos o icnofosil de Chubutolithes gaimanensis
procedentes de Gaiman. El aspecto en
vida, seguramente fue semejante a las avispas de
barro actuales. |
|
Insecto, Hymenoptera.
Se trata de una avispa fósil, conocida
principalmente por la presencia de nidos (icnofosil) en la formación
Sarmiento, en la localidad de Gaiman, Provincia de Chubut, durante
el Eoceno y Oligoceno de la Patagonia. Los sedimentos tienen grandes
espesores de ceniza volcánica aplanada que formaban una vasta
llanura con poco relieve y se cruzaron esporádicamente por los
lechos de los ríos. Este paisaje, similar a la sabana africana, y
fue habitado por diferentes mamíferos y reptiles hoy extintos. Es
posible observar en la formación, abundantes madrigueras de
invertebrados. Encontramos la formación de varios paleosuelos, con
las raíces, nódulos de carbonato y nidos de Chubutolithes
gaimanensis. Estos nidos de barro hechos por avispas, habitaban la
zona hace 39 millones de ańos. Los nidos muestran diferentes capas
de barro utilizadas para construir las paredes del mismo, formando
parte del extrańo ecosistema del Oligoceno. También se conocen
cámaras pupales fósiles de coleópteros, es decir, escarabajos.
Argyrocetus patagonicus. Lydekker, 1894.
|
 |
|
 |
|
Cráneo y mandíbula del Oligoceno tardío - Mioceno temprano
de Chubut. Museo de la Plata. |
|
Aspecto de un integrante de la familia
Eurhinodelphinidae (*). |
Mamífero
Cetáceo.
Eran unos mamíferos cetaceos de la familia extinta
Eurhinodelphinida, unos odontocetos que vivieron del Oligoceno hasta
el Mioceno. Argyrocetus patagonicus, es el miembro más
primitivo de esta familia de cetáceos. Los miembros de familia
poseían una prolongación de la mandíbula superior con apariencia
similar a un pez espada, la cual pudo haber utilizado para golpear o
apuńalar a sus presas o depredadores. De alimentación carnívora y
oportunista. Pudo llegar a medir unos 2 metros de largo y tener un
aspecto semejante a los delfines. La historia evolutiva de los
cetáceos estuvo marcada por al menos tres radiaciones adaptativas.
La segunda radiación (Oligoceno-Mioceno Temprano) marcó la aparición
de los Neocetos (Odontocetos y Mysticetos), y estuvo principalmente
influenciada por la apertura del Océano Austral y cambios en el
ecosistema oceánico y su productividad. Algunos de estos materiales
más modernos, provienen del Oligoceno tardío - Mioceno temprano en
la Formación Gaiman y con una antigüedad de 20 millones de ańos
antes del presente. Tenían una compleja estructura auditiva, que le
permitió desarrollar el sistema de detección través de un
ultrasonido, como las ballenas y delfines actuales. Esto le permitió
emitir sonidos para detectar el tamańo, forma, distancia y velocidad
de un objeto.
Saurodelphis argentinus. Burmeister,1871.
|
 |
 |
|
Mandíbula en el Museo de La
Plata. Mioceno de Paraná. |
Aspecto de un Delfín de
Rió. (*) |
Mamífero
Cetáceo.
Se trata de un mamífero marino odontoceto, es decir,
un cetáceo con dientes como el Cachalote y el Delfín, encontrados en
sedimentos de origen marino depositados durante el Oligoceno
inferior de Patagonia. El mismo comprende de un cráneo muy pequeńo y
rostro extremadamente delgado y alargado, el cual guarda un parecido
muy particular con el Delfín del Ganges, de la costa India. Presenta
ramas mandibulares soldadas en casi todo su largo, formando un hueso
largo y angosto. El origen de los Cetáceos es un misterio, ya que
aparecen súbitamente en la base del Terciario sin mucha evidencia
fósil de sus antecesores, aunque en los últimos ańos se han hallado
en distintas partes del mundo, restos de ballenas y delfines con
rastros de patas traseras, las cuales, evidencian un antecesor
continental y de hábitos carniceros. En el examen de su aparato
dentario, muestra numerosos dientes simples y cónicos, como los
reptiles y su cráneo demuestra cierto parecido a estos animales.
Sufrieron la modificación de sus miembros anteriores, pasando de
fuertes brazos a aletas especializadas para grandes impulsos y la
adaptación de todos los órganos para el medio acuático. Seguramente
se alimentó de peces y organismos menores, como los crustáceos.
|
Ribodon limbatus.
Ameghino,
1883. |
| iboron Riboron Riboron Riboron Riboron Riboron Riboron Riboron Riboron Riboron Riboron Riboron Riboron Riboron Riboron |
|
Esqueletos de Riboron limbatus y reconstrucción en vivo del sirenido fósil de
Patagonia y Paraná, Argentina. (*) |
Mamífero
Sirenido.
Es una especie extinta de
sirénidos, que vivieron en America del Sur.El material que hace
referencia Ameghino, corresponde al Piso Mesopotámico de la
formación Patagónica recogidos en las barrancas del Paraná por el
Profesor Pedro Scalabrini. Los sirénidos, o vacas marinas, son el
único grupo de mamíferos que se han convertido en herbívoros
acuáticos completamente adaptados. En la actualidad, están
representados por tres especies de manatíes (Trichechus) y una sola
especie de dugongos (Dugong dugon). Todos tienen cuerpos bulbosos,
miembros anteriores modificados en forma de aletas, carecen de
extremidades posteriores y tienen una cola aplanada en sentido
horizontal, como las ballenas, que utilizan para impulsarse a través
del agua a un ritmo mesurado. Los sirénidos se conocen desde el
comienzo del Eoceno en Hungría. Su evolución es misteriosa, pero
muchos paleontólogos creen que es posible que compartan un
antepasado con los elefantes. Durante todo el Eoceno, el clima fue
bastante templado y en las aguas tropicales y poco profundas del
Mediterráneo y el Caribe abundaban los prados vastos cubiertos de
hierbas marinas, el principal alimento de los sirénidos marinos. Por
su aparato dentario se acercan a los Notoungulados, pero su
estructura esqueletaría es muy distinta a cualquier otro mamífero
conocido. Se conocen restos fósiles en el Eoceno de Europa y África.
Ribodon tiene su mayor parecido es con el género viviente Manatus,
el cual vive en las desembocaduras de ríos de África occidental y
América oriental, y en la costa marítima oriental en Sudamérica. Los
Lamantiles no se alejan de la zona litoral. La presencia de los
mismos en las opuestas orillas del atlántico, indica claramente que
pasaron de África a Sudamérica emigrando a lo lardo de la costa
desaparecida, por sobre la cual pasaron los mamíferos terrestres,
por el puente Oligoceno.
Carcharodon
megalodon. Agassiz, 1843.
|
 |
 |
 |
|
Diente de
Carcharodon megalodon. (*) |
Mandíbula de
Megalodon recreada a tamańo natural en el Museo
Paleontológico de Bariloche. |
Reconstrucción en vivo (*) |
Pez Selaceo.
Es un enorme tiburón prehistórico que habito
en los antiguos mares del territorio Argentina y distintas partes
del mundo. Poseían un esqueleto cartilaginoso, el cual rara vez se
preserva, pudiendo agregar que los hallazgos de este tipo de
estructura se pueden contar con los dedos de las manos. La única
parte del esqueleto que podemos hallar perfectamente preservado, son
los dientes con esmalte, los cuales varían desde 2 a 20 centímetros.
Sus raros esqueletos se distinguen claramente del gran número de
especies de peces teleósteos (óseos) por su esqueleto cartilaginoso.
Se estima que el tamańo adulto de Carcharodon megalodon, llego a
tener entre 16 y 18 metros de largo. Los tiburones son peces
versátiles y tienen sentidos muy agudos; muchas de sus especies son
capaces de cazar y devorar a casi cualquier otro animal marino,
tanto en mares profundos como en mares poco profundos. Estos dos
rasgos explican su larga historia evolutiva; muchas de las especies
de tiburones de nuestros días son muy similares a especies que
nadaban en los mares del cretácico, hace más de 100 millones de
ańos, o durante el Terciario. Su alimentación estaba constituida por
grandes reptiles marinos de la época, peces y mamíferos marinos
primitivos. Un sitio muy conocido en nuestro país donde se han
hallado restos dentarios de la especie, es Cerro Gualicho en la
Provincia de Rio Negro y en la Formación Paraná, en Entre Ríos, en
sedimentos más recientes del Mioceno.
Archeogeryon
fuegianus. d'Orbigny, 1842.
|
Archaeogeryon peruviensis, Oligoceno superior y Mioceno de
la Provincia de Santa
Cruz, Argentina. (*) El segundo ejemplar
Archeogeryon fuegianus
se exhibe en el
Museo Paleontológico Egidio Feruglio de Trelew (MEF): |
Invertebrado Crustáceo.
Se trata una especie de crustáceo, un
cangrejo marino muy abundante en el registro fosilífero, aunque
generalmente solo se hallan pequeńos fragmentos, y rara vez formas
completas. Estos vivían en los antiguos mares que se hallaban sobre
los actuales sedimentos de Patagonia. Poseen un cuerpo similar a
las especies vivientes, formado por reducción del abdomen y el
ensanchamiento y aumento de tamańo de la parte anterior (delantera)
del cuerpo. El abdomen sirve como bolsa de cría para los huevos, que
en el hembra es más notorio por su tamańo. El cuerpo está más o
menos cubierto por un caparazón quitinoso, con una cubierta cerúlea.
El cuerpo segmentado del cangrejo tiene varios pares de apéndices,
de los cuales cinco suelen servir para la locomoción y dos hacen las
veces de antenas sensoriales. Las patas delanteras estaban equipadas
con pinzas que le servían para alimentarse, defenderse y realizar
exhibiciones rituales de apareamiento. Los cangrejos son capaces de
tolerar cambios en su medio ambiente, los que les permite prosperar
en hábitats muy hostiles, por ello se lo considera junto a los
artrópodos como formas exitosas y primitivas. Sus hábitos
alimentarios son variados. Algunos fósiles de
Archeogeryon fuegianus fueron
hallados muy completos recientemente en los afloramientos geológicos
del Oligoceno de Península de Valdes en la Provincia de Chubut, como
así también en sedimentos de Caleta Oliva en la Provincia de Santa
Cruz.
Otros
Vertebrados del Oligoceno Argentino:
Mamíferos: Proadianthus, Argyrohippus, Cochilius,
Ameghinotherium,
Diochotichus, y Proedrium.
Peces:
Myliobatis sp., Isurus sp., Lamna sp., Odontaspis sp., Hemipristis
sp., Galeocerdo sp.,Carcharodón sp.
(*) Imagen de autor u origen desconocido.
Ver sobre nuestras
Políticas de uso de imágenes.
|
Bibliografía Utilizada.
AMEGHINO, F. 1888. Rápidas
diagnosis de mamíferos fósiles nuevos de la República
Argentina. Buenos Aires, Obras Completas, 5:471-480.
AMEGHINO, F. 1889.
Contribución al conocimiento de los mamíferos fósiles de la
República Argentina. Actas de la Academia Nacional de
Ciencias de Córdoba, 6:1-1027.
Ameghino, F. 1887. Enumeración sistemática de las especies de
mamíferos fósiles coleccionados por Carlos Ameghino en los terrenos
eocenos de Patagonia austral y depositados en el museo de La Plata.
Boletín del Museo de La Plata 1: 1-26.
Bargo, S.; Reguero, M.A.
1989. El primer registro de un mamífero fósil en el extremo
septentrional de Chile. Ameghiniana, Vol. 26, p. 239.
Bellosi, E. 1995.
Paleogeografía y cambios ambientales de la Patagonia Central
durante el Terciario medio. Boletín de Informaciones
Petroleras (BIP), Vol. 44, p. 50-83.
Benton, Michael. (1992).
Dinosaurios y otros animales prehistóricos. Ediciones
Lrousse Argentina S.AI.C.
Bravard, A. (1858). Monografía de los
terrenos marinos terciarios del Paraná. Imprenta del Registro
Oficial 107 pp. Paraná. (Reimpresión del Congreso de la Nación 1995)
Bond,
M.; López,; G.M.; Reguero, M.A.; Scillato-Yané, G.J.; Vucetich, M.G.
1998. Los mamíferos de la Formación Fray Bentos (Edad Deseadense,
Oligoceno superior?) de las provincias de Corrientes y Entre Ríos,
Argentina. Asociación Paleontológica Argentina, Publicación
Especial, Vol. 5, p. 41-50.
Bond, M.; Vucetich, M.G.; Pascual, R.1984.
Un nuevo Notoungulata de la Formación
Lumbrera (Eoceno) de la Provincia de Salta, Argentina. In
Jornadas Argentinas de Paleontología de Vertebrados, No. 1,
Resúmenes, 20 p. La Plata, Argentina.
Candela, A.M.; Morrone, J.J. 2003.
Biogeografía de puercoespines neotropicales (Rodentia,
Hystricognathi): integrando datos fósiles y actuales a
través de un enfoque panbiogeográfico. Ameghiniana, Vol. 40,
p. 361-378.
Cerdeńo, E. y Montalvo, C.I. 2001. Los Mesotheriinae (Mesotheriidae,
Notoungulata) del Mioceno superior de La Pampa, Argentina. Revista
Espańola de Paleontología 16: 63-75.
Barry Cox, Colin Harrison, R.J.G.
Savage, Brian Gardiner. 1999. The Simon & Schuster Encyclopedia of
Dinosaurs and Prehistoric Creatures: A Visual Who's Who of
Prehistoric Life.
Feruglio, E. 1949.
Descripción geológica de la Patagonia. Ministerio de
Industria y Comercio de la Nación, Dirección General de
Yacimientos Petrolíferos Fiscales, Vol. 2, p. 1-349.
Buenos Aires.
Flynn, J.J.; Wyss, A.R.;
Croft, D.A.; Charrier, R. 2003. The Tinguiririca Fauna,
Chile: biochronology, paleoecology, biogeography, and a new
earliest Oligocene South American Land Mammal 'Age'.
Palaeogeography, Palaeoclimatology, Palaeoecology, Vol. 195,
No. 3-4, p. 229-259.
Forster Cooper, C. (1962). Giant rhinoceros from
Loping, Yunnan, and discussion on the taxonomic characters of
Indricotherium grangeri. Vert. Pal. Asiat. 6 (1), pp. 57-71.
Kay, R. F., Campbell, V. M., Rossie, J. B., Colbert, M. W.
and Rowe, T. B. (2004), Olfactory fossa of Tremacebus
harringtoni (platyrrhini, early Miocene, Sacanana,
Argentina): Implications for activity pattern. The
Anatomical Record Part A: Discoveries in Molecular, Cellular,
and Evolutionary Biology, 281A: 1157–1172.
MacFadden, B.J.; Frailey,
C.D. 1984. Pyrotherium, a large enigmatic ungulate (Mammalia,
incertae sedis) from the Deseadan (Oligocene) of Salla,
Bolivia.
Palaeontology, Vol. 27, p. 867-874.
Magnussen Saffer, Mariano (2013). Los principales representantes
de la paleofauna del Oligoceno de la Republica Argentina. Paleo,
Revista Argentina de Paleontología. Boletín Paleontológico. Ańo
11. 82: 11-17.
Magnussen Saffer, Mariano
(2013). Mamíferos carnívoros representativos durante el
Oligoceno. Paleo, Revista Argentina de Paleontología.
Boletín Paleontológico. Ańo 11. 88: 17-22.
Magnussen Saffer, Mariano (2013).
Herbívoros prehistóricos representativos
durante el Oligoceno.
Paleo,
Revista Argentina de Paleontología. Boletín Paleontológico.
Ańo 11. 89: 10-15.
Matsumoto, H. 1922. Revision of
Palćomastodon and Mśritherium. Palćomastodon intermedius, and
Phiomia osborni, new species. American Museum Novitates. Number 51,
November 21.
Matsumoto, H. 1923. A Contribution to
the Knowledge of Mśritherium. Bulletin of the American Museum of
Natural History ; v. 48, article 4. p. 97-140.
Podestá, F. 1899. Un nuevo
fósil Ameghinotherium curuzucuatiensis. La Escuela Positiva,
C.V. p. 1-8.
R. F.
Kay, Fleagle, J.G., Mitchell T.R.T., Colbert M., Bown T.,
Powers D.W. (2008). «The anatomy of Dolichocebus gaimanensis,
a stem platyrrhine monkey from Argentina.». Journal of Human
Evolution 54: 323-382.
Reguero, M.A. 1999. El problema de las relaciones sistemáticas y
filogenéticas de los Typotheria y Hegetotheria (Mammalia,
Notoungulata): análisis de los taxones de Patagonia de la
Edad-mamífero Deseadense (Oligoceno). Tesis Doctoral (Inédito),
Universidad de Buenos Aires, Departamento de Ciencias Biológicas, p.
1-301.
Soria, M.F. (h). 1981. Los Liptoterna del Colhuehuapense (Oligoceno
Tardío) de la Argentina. Revista del Museo Argentino de Ciencias
Naturales “Bernardino Rivadavia”, Serie Paleontología 3: 1-54.
Soria, M.F. (h). 2001. Los Proterotheriidae (Litopterna, Mammalia),
sistemática, origen y filogenia. Monografías del Museo Argentino de
Ciencias Naturales 1: 167 pp.
Vizcaíno SF, Farińa RA. 1997. Diet and locomotion in
Peltephilus: A new view .
Lethaia 30 (1): 79-86
Schlosser Max (1911) "Beiträge zur Kenntnis der
oligozänen Landsäugetiere aus dem Fayum, Ägypten."
Beiträge zur
Paläontologie und Geologie Österreich-Ungarns,
24: 1–167.
Shockey, B.J. 1997.
Toxodontia of Salla, Bolivia (Late Oligocene): Taxonomy,
systematics, and functional morphology.
Ph.D. Dissertation (Unpublished),
University of Florida, p. 1-277.
Ver mas
bibliografía utilizada para hacer la presente Pagina;
AQUÍ.
Ver bibliografía de divulgación sugerida;
AQUÍ. |
|
Periodo
Oligoceno Periodo Oligoceno Periodo Oligoceno Periodo
Oligoceno Periodo Oligoceno Periodo Oligoceno Periodo |
|