El genero Carcharocles desde el Eoceno al
Pleistoceno y su cronología fosilifera.
Por Mariano
Magnussen Saffer. Integrante de Grupo Paleo, Museo Municipal
Punta Hermengo de Miramar y Fundación
Argentavis.
marianomagnussen@yahoo.com.ar. Imágenes ilustrativas de
archivo.
Aviso
Legal
Tomado de;
Magnussen Saffer, Mariano (2014). El genero Carcharocles desde
el Eoceno al Pleistoceno y su cronología fosilífera. Paleo,
Revista Argentina de Paleontología. Boletín Paleontológico. Ańo
XII. 101: 11-16.
Los tiburones
aparecen por primera vez en el registro fósil hace 455 millones
de ańos. Sin embargo, no se vuelven comunes sino hasta el
Devónico, hace unos 400 millones de ańos.
Estudiar la
evolución de estos animales marinos no es fácil ya que al tener
esqueleto de cartílago, no se fosilizan en su totalidad.
Afortunadamente se han encontrado dentículos, dientes y
vértebras fosilizadas que nos permiten conocer los orígenes de
los tiburones.
El tiburón mas
antiguo es el
Antarctilmna
(que significa tiburón lamnoide de la antártica). Se ha
sugerido que este antiguo animal tiene unos 380 millones de ańos
y probablemente vivía en aguas dulces.
 |
Los
tiburones en
sus formas tempranas eran muy pequeńos. Hay pruebas que
sugieren que se parecía más a los peces antes de
evolucionar
a lo que son hoy. Muchos de ellos eran similares a las
anguilas. No tenían un cerebro tan grande como los de
hoy. Más de 2 / 3 del tamańo del cerebro de un
tiburón actual es para los sentidos, sobre todo su capacidad
para oler. Esta es una de las áreas donde los
tiburones se han visto notablemente capaces de
evolucionar
y convertirse en una especie más inteligente. |
Sus
dientes también eran muy diferentes a los de hoy día. En lugar de
tener gran nitidez eran suaves. Sin embargo, estos primeros
antepasados definitivamente tenían las filas de dientes sustituibles
como los actuales. Esto podría indicar que los antepasados de los
tiburones se alimentaban de plantas y que algo ocurrió e hizo
imposible la vida vegetal por un tiempo, con el fin de sobrevivir,
empezaron a consumir otros seres vivos,
evolucionaron.
Probablemente no nadaban tan adentro en el océano como lo hacen hoy.
Esto se evidencia por la forma y el tamańo de sus aletas. Es posible
que hayan tenido que evolucionar
de esta manera para desplazarse largas distancias en
busca de alimentos.
Un
tiburón pierde un diente, éste cae y queda sobre el sedimento. El
diente puede desintegrarse (a) o enterrarse en el sedimento (b);
luego los procesos químicos ocurren; aquí puede que el fósil se
disuelva (c), pero también puede éste absorba los minerales del
sedimento o del agua en los espacios vacíos del diente y así
fosilizarse (d); éste proceso dura en total miles de ańos. El diente
adquiere diferentes colores dependiendo de los minerales que
absorbió. Si la roca es erosionada, después de muchos ańos, es
posible que encontremos éste diente fósil.
Otodus obliquus
es una especie extinta de elasmobranquio lamniforme de la familia
Otodontidae, de gran tamańo, que vivió durante el Paleoceno y el
Eoceno, hace entre 45 y 55 millones de ańos.
 |
Otodus
obliquus
es, según muchos paleontólogos, el ancestro original del más
grande tiburón depredador que ha existido, Carcharodon
megalodon. Hay, sin embargo, otra escuela de pensamiento
que sugiere que C. megalodon fue en realidad
descendiente de Carcharodon orientalis (también
conocido como Paleocarcharodon orientalis) y este
sería el predecesor de los modernos tiburones blancos
Carcharodon carcharias. |
Aunque el argumento de ambos puntos de vista es atractivo, la
evidencia muestra suficientes debilidades para justificar la
precaución en la clasificación de C. megalodon en el mismo
género que el tiburón blanco moderno (es decir, Carcharodon)
y, por lo tanto, ha sido reclasificado recientemente, para reflejar
las pruebas de montaje que Otodus obliquus probablemente fue
el antepasado de C. megalodon.
Nuestra atención, sobre tiburones, se basa en el genero
Carcharocles, cuyos regiros fosilíferos ocupan las capas
sedimentarias desde el Eoceno al Pleistoceno, con formas simples y
corpulentas a terribles monstruos de varias toneladas, representado
por cuatro especies fundamentales.
Carcharocles
auriculatus.
Jordan,
1923.
Carcharocles
auriculatus es una especie extinta de tiburón incluida en el género
Carcharocles, cercanamente relacionada con el mejor conocido
Carcharocles megalodon.
 |
Como ocurre
con muchos tiburones extintos, esta especie es conocida a
partir de dientes fósiles y algunos centros de vértebra. El
esqueleto de los tiburones está compuesto de cartílago y no
de hueso, material que raramente se fosiliza.
La longitud
de los dientes de C.auriculatus es relativamente
grande - de 25 a 114 milímetros. Sin embargo, es menor que
la de C. megalodon y Carcharocles angustidens. |
Muchos de los
dientes de C. auriculatus provienen de Carolina del Sur,
Estados Unidos. Sin embargo, se conocen hallazgos adicionales de
dientes del Eoceno en la meseta Khouribga, en Marruecos.
Carcharocles auriculatus es el miembro más primitivo del género
Carcharocles.
Carcharocles
angustidens. Agassiz, 1843
Carcharocles
angustidens es una especie de tiburón megadentado extinto, que vivió
durante los períodos Oligoceno y Mioceno entre hace aproximadamente
35 a 22 millones de ańos. Se cree que este tiburón es un pariente
cercano del más conocido de los tiburones megadentados, C.
megalodon. Sin embargo, así como en el caso de C. megalodon,
la clasificación de esta especie es materia de debate.
 |
Los restos
de C. angustidens por lo general están en muy mal
estado de preservación. A la fecha, el espécimen mejor
preservado de esta especie ha sido excavado en Nueva
Zelanda, el cual abarca 165 dientes en asociación y cerca de
35 centros de vértebras asociados. Este espécimen data de
hace cerca de 26 millones de ańos. Los dientes de C.
angustidens son notables por sus coronas triangulares y
sus pequeńas cúspides laterales que están totalmente
aserradas. Los bordes aserrados son muy afilados y muy
pronunciados. |
C. angustidens era una especie con una amplia distribución
geográfica con fósiles hallados en Norteamérica, Suramérica, Europa,
África, Nueva Zelanda, Japón, Australia, y Malta.
Igual que en otros tiburones megadentados, los fósiles de C.
angustidens indican que era considerablemente mayor que el
actual gran tiburón blanco, Carcharodon carcharias. Se estima
que el espécimen bien preservado de Nueva Zelanda medía 9.3 metros
de largo. Este ejemplar tenía dientes que miden más de 9.87
centímetros en altura diagonal, y centros de vértebras de cerca de
1.10 centímetros de diámetro. Sin embargo, existen reportes de
fósiles aún mayores de C. angustidens.
Carcharocles
chubutensis.
Es una especie
extinta de tiburón megadentado que vivió durante los períodos
Oligoceno, Mioceno y Plioceno, entre hace 28 a 5 millones de ańos.
Este tiburón es considerado como unpariente cercano de otro tiburón
megadentado prehistórico, el famoso C. megalodon. Sin
embargo, como ocurre con C. megalodon, la clasificación de
esta especie está en discusión.
 |
Esta
especie es conocida a partir de dientes fósiles y algunos
centros de vértebra. El esqueleto de los tiburones se
compone de cartílago y no de hueso, y el cartílago raramente
logra fosilizarse. Por lo tanto, los fósiles de C.
chubutensis están generalmente mal preservados. Aunque
los dientes de C. chubutensis son morfológicamente
similares a los de C. megalodon, estos son
relativamente delgados con una corona curvada, y con la
presencia de talones laterales apenas aserrados. Los fósiles
de esta especie se han hallado en Norteamérica, Sudamérica,
Cuba, Puerto Rico, África, y Europa. |
C. chubutensis
era mayor que C. angustidens. Los dientes de C.
chubutensis se aproximaban a 130 milímetros de altura
perpendicular (longitud diagonal), la cual de acuerdo con el método
de estimación de tamańo propuesto por Gottfried at al, in 1996,
indica que el espécimen tendría una longitud de 12.2 metros.
La investigación
paleontológica sugiere que esta especie puede haber cambiado sus
preferencias de hábitat a través del tiempo, o que pudo haber tenido
la suficiente flexiblilidad en su comportamiento para ocupar
distintos ambientes en distintas épocas.
C. chubutensis
era probablemente un superdepredador y cazaría generalmente peces,
tortugas marinas, cetáceos (por ejemplo, ballenas), y sirenios.
Carcharodon
megalodon.
Agassiz, 1843
es
una especie extinta de tiburón que vivió aproximadamente entre 28 y
1,5 millones de ańos atrás, durante el Cenozoico (de finales del
Oligoceno hasta principios del Pleistoceno).
La
asignación taxonómica de C. megalodon se ha debatido por
cerca de un siglo, y aún se encuentra en disputa con dos
interpretaciones principales: como Carcharodon megalodon
(bajo la familia Lamnidae) o bien como Carcharocles megalodon
(bajo la familia Otodontidae).
C. megalodon
es considerado como uno de los mayores y más poderosos depredadores
en la historia de los vertebrados. Los estudios sugieren que C.
megalodon lucía en vida como una versión corpulenta del gran
tiburón blanco actual, Carcharodon carcharias, llegando a
alcanzar los 16 metros de longitud total. Los restos fósiles indican
que este tiburón gigante tuvo una distribución cosmopolita, con
áreas de cría en zonas costeras cálidas. C. megalodon
probablemente tuvo una influencia muy importante en la estructura de
las comunidades marinas de su época.
 |
C.
megalodon
es representado en el registro fósil principalmente por
dientes y centros de vértebras. Como en los demás tiburones,
el esqueleto de C. megalodon estaba formado de
cartílago más que de hueso; esto resulta en una pobre
preservación de los especímenes encontrados. Sin embargo,
los restos fósiles de C. megalodon indican que tenía
los centros vertebrales densamente calcificados.
Entre las
especies existentes, se considera que el gran tiburón blanco
es la más análoga a C. megalodon. La carencia de
esqueletos fósiles bien preservados de C. megalodon
ha forzado a los científicos a basarse en la morfología del
gran tiburón blanco para inferir su aspecto y estimar su
tamańo. |
Debido a los restos
fragmentarios, estimar el tamańo de C. megalodon se ha
convertido en un reto. Sin embargo, la comunidad científica reconoce
que C. megalodon superaba en tamańo al tiburón ballena (Rhincodon
typus).
En
1973, un ictiólogo de Hawaii, John E. Randall, presentó un método
para estimar la longitud total del gran tiburón blanco. El método
propuesto es representado en forma de un gráfico trazado, el cual
demuestra la relación entre la altura de la corona (la distancia
vertical de la hoja del diente desde la base de la parte de esmalte
del diente hasta su punta) del mayor de los dientes de la mandíbula
superior del gran tiburón blanco y su longitud total
Estos dientes produjeron una longitud total de cerca de 13 metros.
En 1991, dos expertos en tiburones, Richard Ellis y John E. McCosker,
seńalaron un defecto en el método de Randall. De acuerdo a ellos, la
altura de la corona dental de los tiburones no necesariamente se
incrementa en proporción a la longitud total del animal. Esta
observación llevó a nuevas propuestas para dar con métodos más
exactos para determinar el tamańo del gran tiburón blanco y
especies similares.
 |
En 1996
tres científicos – Michael D. Gottfried, Leonard J. V.
Compagno y S. Curtis Bowman – después de haber realizado una
minuciosa investigación y escrutinio de 73 especímenes del
gran tiburón blanco, propusieron una relación linear
conservadora entre la máxima altura del mayor de los dientes
superiores. El mayor diente de C. megalodon al
alcance de este equipo era un espécimen de segundo diente
anterior superior, cuya máxima altura era de 168 mm. Este
diente había sido descubierto por Compagno en 1993, y
produjo una longitud total estimada en 15.9 metros. Sin
embargo, rumores de dientes mayores de C. megalodon
persistían en esa época. La máxima altura dental para este
método se mide como una línea vertical desde la punta de la
corona dental a la parte baja de los lóbulos de la raíz,
paralelo al largo eje del diente. Dicho de otra forma, la
máxima altura del diente es la altura perpendicular.
|
En
2010, varios investigadores de tiburones – Catalina Pimiento, Dana
J. Ehret, Bruce J. MacFadden y Gordon Hubbell — estimaron la
longitud total de C. megalodon sobre la base del método de
Shimada, produjo una longitud total de 16.8 metros.
El
tema de la extinción de C. megalodon está aún bajo
investigación. Hace cerca de 3.1 millones de ańos, ocurrió un
importante evento geológico — el cierre del paso marítimo de
Centroamérica, el cual causó cambios significativos ambientales y en
las faunas alrededor del mundo. Estos cambios probablemente fueron
responsables de la extinción de C. megalodon.
Bibliografía
Sugerida;
Agassiz, L.
1833-1844. Recherches sur les Poissons Fossiles, V. 1-5.
Imprimerie de Petitpierre, Neuchâtel, Suisse.
Bretton W. Kent (1994). Fossil Sharks of the Chesapeake Bay Region.
Egan Rees &
Boyer, Inc. 146 páginas.
Bruner, John (1997).
«The "Megatooth" shark, Carcharodon megalodon» Mundo Marino Revista
Internacional de Vida Marina.
Consultado el 22-01-2012.
Cione A.L., E.P. Tonni y J.O. San Cristóbal.
A
middle Pleistocene marine transgression in central-eastern
Argentina. Current Research in the Pleistocene 19: 16-18; 2002.
Lambert, Olivier; Giovanni Bianucci, Klaas Post, Christian de Muizon,
Rodolfo Salas-Gismondi, Mario Urbina and Jelle Reumer (1 de julio de
2010). «The giant bite of a new raptorial sperm whale from the
Miocene epoch of Peru». Nature (Peru) 466 (7302): pp. 105–108.
Gottfried M.D., Fordyce R.E (2001). «An Associated Specimen of
Carcharodon angustidens (Chondrichthyes, Lamnidae) from the Late
Oligocene of New Zealand, with comments on Carcharodon
Interrelationships». Journal of Vertebrate Paleontology 21 (4):
pp. 730–739.
Glikman, L.S., 1980.. Evolution of Cretaceous and Caenozoic Lamnoid
Sharks:3-247, pls.1-33. Moscow.
Jordan, D.S. & Hannibal, H., 1923. Fossil Sharks and Rays of the
Pacific Slope of North America. Bulletin of the Southern California
Academy of Sciences, 22:27-63, plates 1-9.
Marsili, Stefano; Giorgio Carnevale, Ermanno Danesea, Giovanni
Bianuccia, and Walter Landinia (March 2007).
«Early Miocene
vertebrates from Montagna della Maiella, Italy». Annales de
Paléontologie (Italy: Elsevier) 93 (1): pp. 27–66.
Rudwick, M. J. S.
(1987) [1976]. El significado de los fósiles. Episodios de la
Historia de la Paleontología. Ciencias de la Naturaleza. Madrid:
Hermann Blume. pp. 347.
Wroe, S.; Huber, D.
R. ; Lowry, M. ; McHenry, C. ; Moreno, K. ; Clausen, P. ; Ferrara,
T. L. ; Cunningham, E. ; Dean, M. N. ; Summers, A. P. (2008).
«Three-dimensional computer analysis of white shark jaw mechanics:
how hard can a great white bite?».
Journal of
Zoology 276
(4): pp. 336–342.
Los dinosaurios de la Argentina.
Por Leonardo Salgado,
Museo de Geología y Paleontología,
Universidad Nacional del Comahue,
lsalgado@uncoma.edu.ar
y Ricardo Pasquali,
Universidad Tecnológica
Nacional, Facultad Regional Haedo,
rcpasquali@yahoo.com.
Articulo publicado originalmente en
www.museosargentino.com.ar Imágenes colocadas por PaleoArgentina.
Aviso
Legal
Una reseńa sobre las principales
especies conocidas y su importancia paleontológica.
Los dinosaurios
evolucionaron a partir de algún pequeńo reptil que vivió hace unos
230 millones de ańos, en una época que los geólogos y paleontólogos
denominan Triásico tardío.
Una de las especies que guarda mayor
relación con los dinosaurios es el Lagosuchus talampayensis.
Los
restos de este reptil fueron hallados en la provincia de La Rioja en
1964, durante una expedición organizada por el paleontólogo Alfred
Romer, de la Universidad de Harvard, con la colaboración del Museo
de La Plata. En 1975, José Bonaparte, que entonces se desempeńaba
como investigador del CONICET en la Fundación Miguel Lillo de
Tucumán, descubrió ejemplares más completos y publicó una detallada
descripción de la casi totalidad del esqueleto de este animal, en la
que se destacaban varios caracteres en común con los dinosaurios.
Casi veinte ańos más tarde, el paleontólogo estadounidense Paul
Sereno, de la Universidad de Chicago, junto con Andrea Arcucci, de
la Universidad de La Rioja, propusieron que el espécimen estudiado
por Bonaparte como Lagosuchus talampayensis correspondería en
realidad a un nuevo género y especie, a la que denominaron
Marasuchus lilloensis. De todos modos, estuvieron absolutamente de
acuerdo en que ese animal estaba, desde un punto de vista evolutivo,
a un paso de distancia de los dinosaurios.
Otro reptil que guarda relación con los
primeros dinosaurios es el Pseudolagosuchus major, descripto por
Arcucci en 1987. Bonaparte considera que el Pseudolagosuchus major
es en realidad un adulto del Lagosuchus talampayensis. Tanto el Lagosuchus talampayensis, como el Marasuchus lilloensis y el
Pseudolagosuchus major, se descubrieron en unos estratos geológicos
conocidos como Formación Los Chańares, del Triásico medio de La
Rioja.
Una de las características más
llamativas del cráneo de los dinosaurios es la presencia de dos
aberturas ubicadas detrás de cada uno de los ojos, una encima de la
otra, que permitirían a los músculos de la mandíbula aumentar de
volumen cuando se contraían durante la masticación, y de una
abertura delante de cada ojo que podía haber contenido una glándula.
Esta particularidad del cráneo hace que se los clasifique como
arcosaurios –que en griego significa reptiles dominantes– junto con
sus parientes los los pterosaurios –reptiles voladores–, los
cocodrilos, y una gran variedad de formas extinguidas, entre ellas,
el Pseudolagosuchus y el Marasuchus.
Todos los dinosaurios se caracterizan
por la posición erecta de las extremidades –parecida a la que se
encuentra en los mamíferos– por la cual sujetan al cuerpo desde
abajo, manteniéndolo apartado del suelo. Cada una de las
extremidades se mueve en un plano más o menos vertical. Esta postura
contrasta con la posición casi horizontal de las extremidades de la
mayoría de los demás reptiles, los cuales descansan con el vientre
en el suelo y los brazos y muslos proyectados lateralmente desde el
cuerpo. En realidad, esta particular disposición de los miembros
estaba ya presente en los pequeńos precursores Lagosuchus y
Marasuchus. żCuáles son, entonces, las características propias de
los dinosaurios, aquellas que permiten distinguirlos de todos los
demás reptiles? Las más destacadas se aprecian en el tarso, que
facilitaban el hábito digitígrado de los dinosaurios, y en el
acetábulo, la cavidad de la cadera en la que se inserta la cabeza
del fémur. El acetábulo de los dinosaurios tiene un borde superior
muy desarrollado y un gran orificio en el fondo. En los restantes
reptiles, en cambio, el acetábulo no presenta ninguno de estos
rasgos. Otra característica de los dinosaurios es el pescuezo con
forma de S.
La diferenciación de los dinosaurios
Al poco tiempo de aparecer, los
dinosaurios se diferenciaron en dos grandes grupos:
ornitisquios y saurisquios.
En la pelvis de los ornitisquios –en
griego, cadera de ave– el pubis estaba orientado hacia atrás y
abajo, como en las aves actuales. En la de los saurisquios –en
griego, cadera de lagarto– en cambio, el pubis estaba orientado
hacia abajo y adelante, manteniendo la disposición que se observa en
otros grupos más primitivos.
En 1967, Rodolfo Casamiquela describió
en Ameghiniana, la revista de la Asociación Paleontológica
Argentina, al dinosaurio ornitisquio más antiguo conocido, al que
llamó Pisanosaurus mertii. Los huesos de este dinosaurio del
Triásico tardío, descubierto en la Hoyada de Las Lajas, en la
provincia de La Rioja, habían sido exhumados por Bonaparte, el
paleobotánico Rafael Herbst, Galileo Scaglia –director del Museo
Municipal de Mar del Plata y Martín Vince en 1962. Los sedimentos en
los que se realizó este descubrimiento pertenecen a la llamada
Formación Ischigualasto, depositados hace unos 230 millones de ańos.
El Pisanosaurus mertii era un dinosaurio herbívoro –como lo eran
todos los ornitisquios– que medía 1 metro de largo.
Los saurisquios más antiguos de la
Argentina –y entre los más antiguos del mundo– son el
Herrerasaurus
ischigualastensis, descripto por Osvaldo Reig, de la Universidad de
Buenos Aires, en 1963, y el Eoraptor lunensis, descripto en 1993 por
Paul Sereno, Catherine Foster y Raymond Rogers, de la Universidad de
Chicago, junto con Alfredo Monetta, del Museo de Ciencias Naturales
de la Universidad Nacional de San Juan. Ambos dinosaurios carnívoros
fueron hallados en Ischigualasto, provincia de San Juan, en
sedimentos de la Formación Ischigualasto, y por lo tanto tienen la
misma antigüedad que el ornitisquio Pisanosaurus mertii.
El Herrerasaurus fue descubierto por la
expedición de la Universidad Nacional de Tucumán dirigida por Reig.
El hallazgo lo realizó el guía Victorino Heredia, habitante del
lugar, a quien Reig le dedicó el nombre genérico del dinosaurio.
Este primitivo saurisquio medía unos 4 metros de largo por 2 metros
de alto hasta el cráneo. Caminaba en postura bípeda y tenía los
miembros anteriores bastante reducidos. Bonaparte sintetiza las
características de este dinosaurio diciendo que se trata de un
saurisquio muy arcaico con rasgos primitivos en los miembros
posteriores, sacro e ileon, pero con caracteres derivados en las
vértebras y pubis.
El Eoraptor fue descripto a partir de un
pequeńo espécimen juvenil descubierto por el geólogo Roberto
Martínez, del Museo de Ciencias Naturales de la Universidad Nacional
de San Juan, integrante de la expedición organizada por la
Universidad de Chicago y la Universidad Nacional de San Juan. Este
dinosaurio medía 1 metro de largo y, como el Herrerasaurus,
caminaba en posición bípeda. De acuerdo con Bonaparte, el Eoraptor
probablemente es un género primitivo de la familia de los
herrerasáuridos, a la cual pertenece el Herrerasaurus.
Los dinosaurios con cadera de ave
En Argentina, los restos de ornitisquios
son aún escasos, si se tiene en cuenta el gran registros con que
cuentan los saurisquios. Algunos fueron posiblemente autóctonos y
otros ingresaron a América del Sur como consecuencia de la aparición
de cadenas de islas transitorias que permitieron intercambios
faunísticos ocasionales con América del Norte durante el Cretácico
tardío. Entre estos ornitisquios que se conocen en nuestro país
tenemos a los dinosaurios "pico de pato", o hadrosaurios; los
pequeńos iguanodontes; un posible dinosaurio con cuernos, o
ceratópsio, y un dinosaurio con armadura, o anquilosaurio.
|
 |
Los hadrosaurios presentaban algunas
adaptaciones extraordinarias, sobre todo aquellas relacionadas con
la herbivoría. Tenían un aspecto muy diferente unos de otros y
presentaban distintos tipos de protuberancias y crestas óseas sobre
la cabeza. El rasgo distintivo de estos dinosaurios es la forma en
que se alargaba la parte anterior de la cara para formar un hocico
ancho y aplanado, con un pico desprovisto de dientes, que por su
aspecto recuerda al de un pato. Para fines del Cretácico, los
hadrosaurios se habían extendido por todas las regiones del
Hemisferio Norte y habían ingresado a América del Sur.
|
El hadrosaurio de la Argentina mejor
documentado es el Kritosaurus australis. Este dinosaurio, que medía
hasta 5 metros de largo, fue descubierto en la provincia de Río
Negro, en
sedimentos de la
Formación Los Alamitos, depositados hace unos 70 millones de ańos.
Fue descripto en 1984 por Bonaparte, M. R. Franchi, Jaime Powell y
Eliseo Sepúlveda.
El Kritosaurus australis comparte muchas
características con el Kritosaurus navajovius, de Nueva México. La
comparación de ambos dinosaurios sugiere que los hadrosaurios
colonizaron América del Sur, provenientes de América del Norte, en
un tiempo relativamente breve.
Los paleontólogos Rodolfo Coria
–director del Museo “Carmen Funes” de Plaza Huincul– y Leonardo
Salgado –investigador de la Universidad Nacional del Comahue–descubrieron
en la localidad rionegrina de Cinco Saltos numerosos representantes
de un grupo de pequeńos ornitisquios bípedos, relacionados con los
hadrosaurios, que vivieron al igual que estos últimos en el
Cretácico tardío. Los dos paleontólogos hallaron una docena de
especímenes juveniles y adultos de estas criaturas describiéndolas
en 1996 con el nombre de Gasparinisaura cincosaltensis, en
reconocimiento a la doctora Zulma Gasparini, especialista en
reptiles mesozoicos del Museo de La Plata. Entre los materiales más
importantes que se hallaron figura un cráneo diminuto, en excelente
estado de preservación. Se puede ver en él, una apretada hilera de
dientes, de características diferentes a los de los hadrosáuridos.
En 1918, el geólogo Augusto Tapia
–de la entonces Dirección Nacional de Minería– anunció el hallazgo
de una mandíbula incompleta de un dinosaurio ceratopsio –grupo que
incluye a dinosaurios herbívoros con cuernos, como el Triceratops de
América del Norte– descubierto en las cercanías del lago Colhué-Huapi,
en la provincia de Chubut. Esta mandíbula, que representó el primer
hallazgo de dinosaurios ornitisquios en América del Sur, fue
estudiada en 1929 por el paleontólogo alemán Friederich von Huene,
de la Universidad de Tübingen, que fuera contratado por Luis María
Torres, director del Museo de La Plata. Lo curioso del hallazgo de
Tapia –hoy extraviado–, es que jamás se ha vuelto a hallar otro
resto de este grupo de dinosaurios.
En 1996, Salgado y Coria anunciaron en
Ameghiniana el primer hallazgo de un anquilosaurio en América del
Sur. Los anquilosaurios eran dinosaurios ornitisquios macizos,
cuadrúpedos y herbívoros en los cuales el cuello, el dorso, los
flancos y la cola estaban recubiertos por completo de un mosaico de
placas dérmicas óseas planas, cubiertas por púas y protuberancias de
distintos tipos. El espécimen, un pequeńo fémur derecho, fue
descubierto en Salitral Moreno, al sur de General Roca, provincia de
Río Negro, por Juan Sepúlveda, del Museo Provincial “Carlos Ameghino”
de Cipolletti. Este dinosaurio acorazado vivió en el Cretácico
tardío. En sucesivas campańas, los mismos investigadores pudieron
hallar más material: vértebras, placas y un diente.
Los primeros dinosaurios con cadera de
lagarto
En el Triásico tardío los dinosaurios
saurisquios –de cadera de lagarto– se habían diferenciado en dos
grupos: Unos, los terópodos, mantuvieron el andar bípedo de sus
precursores; el otro grupo, los sauropodomorfos, adoptaron en cambio
el cuadrupedalismo. Los primeros fueron mayormente depredadores; los
segundos, herbívoros.
Los terópodos del Triásico mejor
conocidos son el Herrerasaurus ischigualastensis y el Eoraptor
lunensis. Este último es muy importante por haber retenido algunos
caracteres primitivos que en el resto de los dinosaurios se han
modificado, por ejemplo, el extremo no expandido del pubis, uno de
los huesos de la cadera.
Los sauropodomorfos del Triásico, los
prosaurópodos, eran de talla moderada a grande y se extinguieron a
principios del Jurásico. Están representados en la Argentina por el
Coloradisaurus brevis, el Mussaurus patagonicus –de la familia de
los plateosáuridos–, el Lessemsaurus sauropoides y el
Riojasaurus
incertus, perteneciente a la familia de los melanorosáuridos. Los
miembros de este último grupo, al que pertenece el Melanosaurus
readi, de Sudáfrica, eran los más grandes de los prosaurópodos.
 |
Tanto el Riojasaurus incertus como el Coloradisaurus brevis y el
Lessemsaurus sauropoides provienen de la
Formación Los Colorados, de la provincia de La Rioja, con una
antigüedad menor que la Formación Ischigualasto. La asociación
faunística documentada en estos estratos, que representan la parte
final del Triásico, estaba dominada por dinosaurios prosaurópodos. El
Riojasaurus incertus medía hasta 7
metros de largo. Fue descubierto por Bonaparte en la Quebrada de los
Jachaleros, La Rioja, quien lo describió en 1969. |
El estudio del cráneo fue realizado por
Bonaparte y José Pumares, quienes
publicaron sus resultados en 1995. Los caracteres de este dinosaurio
sugieren que posiblemente sea el más primitivo de los prosaurópodos. El prosaurópodo Coloradisaurus brevis
mediría unos 3 metros de largo y aproximadamente 1 metro de alto en
posición cuadrúpeda. En posición bípeda la altura de la cabeza
estaría a unos 2 metros. Este dinosaurio es más evolucionado que el
Riojasaurus incertus. Fue descubierto y descripto por Bonaparte en
1978, quién lo incluyó en la familia de los plateosáuridos, a la que
pertenece el Plateosaurus engelhardti, un prosaurópodo del Triásico
tardío descubierto en Europa.
Entre el numeroso material de
dinosaurios de la Formación Los Colorados figura el Lessemsaurus
sauropoides, descubierto por Bonaparte, quien lo describió en 1999.
Se trata de un conjunto de vértebras cervicales y dorsales,
asociados con algunas piezas de las cinturas, que poseen ciertos
rasgos avanzados que se hallan presentes en los saurópodos. Este
último grupo de enormes dinosaurios herbívoros, sin embargo, recién
se diversificará y prosperará a partir del periodo siguiente: el
Jurásico.
Los especímenes del Mussaurus
patagonicus fueron descubiertos por Bonaparte y sus asistentes en la
provincia de Santa Cruz, en sedimentos del Triásico tardío
pertenecientes a la Formación El Tranquilo, depositados hace unos
220 millones de ańos, aproximadamente. De este dinosaurio se conoce
un ejemplar juvenil de unos 16 centímetros de largo, que fue
descripto por Bonaparte y Martín Vince en 1979. El mismo fue
encontrado junto a siete individuos del mismo tamańo y dos huevos,
los que formaban aparentemente parte de un nido. Este hecho indica
que el cuidado de las crías era una conducta que estaba presente en
dinosaurios muy antiguos.
Los dinosaurios del Jurásico
En 1947, el científico espańol Ángel
Cabrera, que para entonces se desempeńaba como jefe de la División
Paleontología de Vertebrados de la Facultad de Ciencias Naturales y
Museo de la Universidad Nacional de La Plata, estudió los restos del
primer saurópodo del Jurásico registrado en América del Sur. El
hallazgo lo había realizado el geólogo de Yacimientos Petrolíferos
Fiscales Tomás Suero en Cerro Negro, Pampa de Agnia, provincia de
Chubut. Este dinosaurio, al que Cabrera nombró como Amygdalodon
patagonicus, pertenece al grupo de los cetiosáuridos (cetiosaurio
significa lagarto ballena). Casamiquela completó su estudio y
publicó sus resultados en 1963. Este hallazgo sirvió para demostrar
que, durante el Jurásico, América del Sur mantenía intercambios de
faunas continentales con otras regiones del mundo, ya que los
cetiosaurios se registraron en el norte de África, Europa y Asia.
Posiblemente el Amygdalodon patagonicus vivió en la parte final del
Jurásico temprano.
La mayor parte de los dinosaurios
jurásicos conocidos de la Argentina provienen del Cerro Cóndor, en
la provincia del Chubut. Los afloramientos del Jurásico medio de
este yacimiento paleontológico corresponden a la Formación Cańadón
Asfalto, y poseen una antigüedad de unos 165 millones de ańos. Esta
localidad fosilífera fue trabajada por Bonaparte y colaboradores
desde la Universidad Nacional de Tucumán y posteriormente desde el
Museo Argentino de Ciencias Naturales “Bernardino Rivadavia”, entre
1976 y 1985.
En el Cerro Cóndor se descubrieron el
terópodo Piatnitzkysaurus floresi y los saurópodos
Patagosaurus
fariasi y Volkheimeria chubutensis.
El primer ejemplar del Piatnitzkysaurus
floresi fue colectado en 1977 y descripto por Bonaparte en 1979.
Este dinosaurio carnívoro medía unos 5 metros de largo y tenía
cavidades neumáticas en las vértebras que, según Bonaparte, habrían
contribuido a la oxigenación de la sangre. Las características de la
dentadura sugieren que este animal no era un feroz predador de
presas grandes, sino de vertebrados relativamente pequeńos. Muestra
similitudes con el Allosaurus
fragilis, un
carnívoro de gran tamańo del Jurásico tardío de América del Norte,
pero es más primitivo. El Piatnitzkysaurus floresi parece ser un
miembro de la misma línea evolutiva que condujo al Allosaurus.
Bonaparte lo considera como un miembro primitivo de la familia de
los allosáuridos.
| |
|
 |
El Patagosaurus fariasi, que llegaba a
medir unos 14 metros de largo, es el dinosaurio más frecuente de
Cerro Cóndor. Fue descripto por Bonaparte en 1979 como perteneciente
al grupo de los cetiosáuridos, la misma familia del Amygdalodon
patagonicus. El descubrimiento de dos adultos y tres juveniles de
diferente tamańo encontrados juntos sugiere que pueden representar
un grupo familiar. La escápula del Patagosaurus fariasi posee una
morfología primitiva del tipo presente en los géneros Cetiosaurus,
del Jurásico medio de Inglaterra, y Barapasaurus, del Jurásico
temprano de la India. La anatomía vertebral de estos tres
dinosaurios es menos derivada que en los saurópodos del Jurásico
tardío, tales como el Diplodocus y el Camarasaurus, de América del
Norte, y el Brachiosaurus, de América del Norte y África. El
Volkheimeria chubutensis es un
saurópodo cetiosáurido descripto por Bonaparte en 1979 a partir de
unas pocas piezas diagnósticas. Es muy similar al Lapparentosaurus
madagascarensis, del Jurásico medio de Madagascar.
|
Recientemente ha sido descripta una
nueva especie de saurópodo de los mismos niveles estratigráficos: el
Tehuelchesaurus benitezii. Lo destacable de esta especie es la
notable similitud que posee con un saurópodo de China, lo que
indicaría una cierta continuidad faunística entre Patagonia y Asia
Oriental durante ese tiempo.
Algunos de los dinosaurios del Jurásico
tardío están representados por huellas en unas areniscas en la
Estancia Laguna Manantiales, en el noreste de Santa Cruz, las que
fueron descubiertas y estudiadas por Casamiquela. En 1964, este
investigador, que entonces se desempeńaba en el Museo de La Plata,
describió huellas fósiles de tres tipos de dinosaurios pequeńos y de
un mamífero primitivo. Uno de los dinosaurios era un cuadrúpedo,
posiblemente ornitisquio, con un pie de solo tres centímetros de
largo, al que Casamiquela denominó Sarmientichnus scagliai.
Otro era
un dinosaurio carnívoro que caminaba en posición bípeda, el Wildeichnus navesi, y el tercero también era un carnívoro bípedo,
pero de mayor tamańo, al que llamó Delatorrichnus goyenechei.
El hecho de que el registro de
dinosaurios del Jurásico tardío sea tan escaso, tanto en nuestro
país como en el resto de América del Sur, impide efectuar
comparaciones adecuadas con faunas bien conocidas de otros puntos
del planeta, como África y América del Norte, en donde el registro
de especies de ese período es excelente.
El Cretácico temprano
Los dinosaurios del Cretácico temprano
de la Patagonia provienen de las Formaciones La Amarga, Rayoso y
Lohan Cura, que corresponden a lapsos del tiempo geológico conocidos
como edades Neocomiano la primera y Aptiano la segunda y tercera, y
del Miembro Candeleros de la Formación Río Limay, que corresponde a
la edad Albiano.
La localidad de La Amarga está ubicada a
unos 70 kilómetros al sur de Zapala, en la provincia del Neuquén, en
la que están expuestas las areniscas que componen la Formación La
Amarga. De estas areniscas, formadas hace unos 130 millones de ańos,
provienen restos de cocodrilos, pterosaurios, mamíferos y
dinosaurios, siendo los mejores conocidos el saurópodo con espinas
Amargasaurus cazaui y el pequeńo terópodo Ligabueino andesi. La
exploración paleontológica de estos estratos fue comenzada por
Bonaparte y colaboradores a partir de las informaciones dadas por el
geólogo Luis Cazau.
El Amargasaurus cazaui era un herbívoro
de unos 8 metros de largo que tenía dos filas de largas espinas que
le recorrían el cuello y el lomo. Bonaparte pudo determinar que el
Amargasaurus era un animal estrechamente emparentado con dos
especies del Jurásico tardío de Tanzania, África oriental,
pertenecientes al género Dicraeosaurus; las tres especies integraban
una misma familia zoológica. En el Dicraeosaurus y en el
Amargasaurus las espinas neurales de las vértebras no eran simples
como en el Patagosaurus, el Volkheimeria u otros saurópodos, sino
que se bifurcaban en la parte superior formando una Y. Estas largas
espinas se extendían por toda la espalda y el cuello, dándole al
animal un aspecto realmente extrańo.
Bonaparte supone que la selección
natural favoreció a estos animales debido a que las enormes espinas
neurales constituían una defensa contra el ataque de los carnívoros.
La persistencia de la presión de selección ejercida por los
depredadores posiblemente condujo a ciertas especies a un desarrollo
extremo de las espinas neurales bifurcadas, como puede verse en el
Amargasaurus. La descripción de este saurópodo la realizó Bonaparte
en 1991 junto con Leonardo Salgado.
El Ligabueino andesi fue descubierto en
1985 por la expedición paleontológica del Museo Argentino de
Ciencias Naturales dirigida por Bonaparte, quien lo describió en
1996. Su pequeńo tamańo sugiere que, posiblemente, se trataba de un
terópodo insectívoro. El tamańo del fémur es de sólo 5 centímetros
de largo y el largo total seguramente no sobrepasaba los 50
centímetros. Representa el registro más antiguo de la familia de los
abelisáuridos, a la que pertenecen también los terópodos Abelisaurus
comahuensis, del Cretácico tardío de Río Negro, y Carnotaurus
sastrei, del Cretácico de Chubut. Este grupo parece ser endémico del
supercontinente de Gondwana, que estaba formado por América del Sur,
África, la India, la Antártida, Australia y Nueva Zelanda. Al igual
que en el Carnotaurus, el pequeńo terópodo de La Amarga posee arcos
neurales extremadamente neumatizados que, de acuerdo con Bonaparte,
podrían ayudar a la respiración en ciertas situaciones de altas
demandas de oxígeno.
La Edad Aptiana, que comprende el lapso
comprendido entre 115 y 108 millones de ańos atrás, está
representada en la Patagonia por las Formaciones Rayoso y Lohan
Cura.
| |
|
 |
La Formación Rayoso aflora en las
cercanías de Agrio del Medio, provincia del Neuquén. De esta
formación geológica proviene el Rayososaurus agrioensis, un
saurópodo descubierto y descripto por Bonaparte en 1995. En la Formación Lohan Cura, en las
cercanías de Picún Leufú, Neuquén, la expedición del Museo Argentino
de Ciencias Naturales descubrió dos saurópodos muy particulares.
Uno, al que Bonaparte llamó Agustinia ligabuei, fue hallado en 1997
por Agustín Martinelli, integrante de la expedición. Este dinosaurio
poseía a lo largo del dorso unas placas de más de sesenta
centímetros de largo dispuestas de a pares. Otro ejemplar, referido
al Agustinia, tiene placas de más de 1 metro de largo.
|
El otro saurópodo, descubierto en 2000
tenía una postura similar a la jirafa, con los miembros anteriores
más largos que los posteriores. Otro dinosaurio con esta postura fue
el braquiosaurio –Brachiosaurus altithorax–, del Jurásico tardío de
América del Norte, que vivió unos 40 millones de ańos antes que el
descubierto en Neuquén.
Otra característica del Agustinia
ligabuei que lo hace excepcional es la presencia de grandes huecos
en las vértebras, lo que hacía que el esqueleto fuera más liviano y
más fuerte que si hubiese sido macizo. Según Bonaparte, este tipo de
estructura era capaz de absorber vibraciones y resistir tensiones,
además de ser flexible.
Del Miembro Candeleros de la Formación
Río Limay proceden los saurópodos Andesaurus delgadoi, descripto por
Calvo y Bonaparte en 1991, Rebbachisaurus tessonei descripto por
Calvo y Salgado en 1995, y el terópodo Giganotosaurus carolinii,
descripto por Coria y Salgado en 1995.
El Andesaurus delgadoi medía unos 15
metros de largo y su cabeza habría estado a varios metros de altura.
Era un primitivo titanosaurio relacionado a los saurópodos
brachiosáuridos –grupo al que pertenece el Brachiosaurus–, y procede
de la zona de El Chocón, Neuquén. Este dinosaurio fue detectado por
Alejandro Delgado, radiólogo del hospital de El Chocón mientras
buceaba en el lago Ezequiel Ramos Mexía. Su extracción se realizó,
en colaboración con el Museo Argentino de Ciencias Naturales, una
vez que el nivel del lago descendió y lo dejó al descubierto.
El Rebbachisaurus tessonei es el
saurópodo más completo hallado en América del Sur hasta el momento.
Medía 17 metros de largo y está relacionado con el Rebbachisaurus
garasbae, del Cretácico temprano de Marruecos, y con el
Diplodocus
longus, del Jurásico tardío de los Estados Unidos. Sin embargo la
especie neuquina es más primitiva que el Diplodocus. Una notable
diferencia entre ambas especies es que las espinas neurales de las
vértebras cervicales y dorsales en el Rebbachisaurus no tienen forma
de "V" como en el resto de los diplodócidos.
Hasta ahora el Rebbachisaurus tessonei
es el único saurópodo en el cual se puede confirmar la existencia de
piedras gástricas. Estas piedras, también llamadas gastrolitos,
cumplirían la función de moler los vegetales en el interior del
estómago, ya que estos dinosaurios no eran capaces de masticar la
comida que ingerían.
Los dos saurópodos que se hallaron en El
Chocón poseen historia evolutivas diferentes. Mientras que el
Andesaurus es parte de un linaje que saurópodos que florecerá en el
Cretácico tardío, el Rebbachisaurus (forma afín al
Rayososaurus)
pertenece a una estirpe que se extinguirá definitivamente a fines
del Cretácico temprano. Al igual que el Giganotosaurus, ambos
saurópodos poseen, en la misma época, parientes cercanos en el
África. Esto sin duda se vincula con la existencia de una conexión
terrestre entre ambos continentes, la que pudo haberse roto recién
en el Cenomaniano.
El Giganotosaurus carolinii, el mayor de
los dinosaurios carnívoros del mundo, fue descubierto en El Chocón,
provincia del Neuquén, por Rubén Carolini. Sus restos fueron
extraídos en 1993 por un equipo de paleontólogos de la Universidad
Nacional del Comahue y del Museo "Carmen Funes" de Plaza Huincul. El
cráneo de este dinosaurio mide aproximadamente 1,80 metros de largo,
mientras que el largo total del animal se estima en unos 16 metros.
El nombre de este gigantesco dinosaurio significa “lagarto gigante
del sur de Carolini”. Debido a que fue hallado en los mismos niveles
que el Giganotosaurus, el saurópodo Rebbachisaurus tessonei
seguramente fue una de las víctimas de este temible predador.
El Carnotaurus sastrei, uno de los más
espectaculares terópodos del mundo, posiblemente vivió durante el
Albiano, 107 a 95 millones de ańos atrás. Es el primer dinosaurio
carnívoro conocido que poseía cuernos y del cual se conserva la
impresión de la piel.
De acuerdo con Bonaparte, que describió
al Carnotaurus sastrei en 1985, las características anatómicas de
este dinosaurio indican un endemismo muy marcado. Una de las tantas
particularidades de este dinosaurio son sus pequeńos miembros
anteriores, muchos más reducidos que los del Tyrannosaurus. La
estructura de las vértebras del cuello del Carnotaurus era muy
especial y parecida a la de las aves, posiblemente con una
musculatura cruzada que permite hacer más movimientos y mucho más
precisos. La presencia de cuernos, sumado a la reducción de los
miembros anteriores, tendría que ver con la destreza muy marcada que
posiblemente tuvo este animal con sus cuernos, para herir o
despanzurrar a otro animal. Evidentemente tenía una precisión muy
grande en los movimientos de la cabeza, que lo sugiere ese tipo de
estructura vertebral del cuello.
Los gigantes del Cretácico tardío
Las enormes vértebras de uno de los
dinosaurios más grandes del mundo, el Argentinosaurus huinculensis,
ocupan casi una sala del Museo "Carmen Funes" de Plaza Huincul,
provincia del Neuquén. Este gigante, que procede del Miembro Huincul
de la Formación Río Limay, pertenece al amplio grupo de los
titanosaurios y vivió hace algo menos de 95 millones de ańos. Sus
restos fueron estudiados por Bonaparte y Coria, quienes publicaron
su descripción en 1993.
Las vértebras del Argentinosaurus
huinculensis, que poseen características anatómicas básicamente
similares a la de otros titanosaurios, miden entre 1,40 y 1,60
metros de altura y 1,30 metros de ancho. Se estima que el
Argentinosaurus poseía un largo de unos 40 metros y su peso superaba
las 100 toneladas.
| |
|
 |
Otro saurópodo titanosaurio muy
particular del Cretácico tardío, aunque algo más reciente que el
Argentinosaurus huinculensis, es el Saltasaurus loricatus, descripto
por Bonaparte y Jaime Powell en 1980. Estos investigadores
estudiaron los restos de cinco especímenes que se hallaron juntos en
la Estancia El Brete, en el sur de la provincia de Salta. Los
sedimentos portadores corresponden a la Formación Lecho, de edad
Maastrichtiana, la última de la era Mesozoica, con una antigüedad de
unos 70 millones de ańos. El Saltasaurus loricatus medía hasta 10
metros de largo y tenía el cuerpo protegido por placas de 10
centímetros de diámetro y pequeńos osículos de 1 centímetro.
|
El Argentinosaurus y el Saltasaurus nos dan una idea del amplio rango
de tamańos que tuvieron los titanosaurios.
También vivió a fines del Cretácico el
Aelosaurus rionegrensis, un titanosáurido descripto por Powell en
1987 cuyo largo total era de unos 15 metros. Este dinosaurio fue
descubierto en la provincia de Río Negro, en sedimentos
pertenecientes a la Formación Los Alamitos. En 1993, Salgado y Coria
describieron nuevos restos del Aelosaurus provenientes de la
localidad rionegrina de Salitral Moreno, entre los cuales había dos
placas dérmicas de unos 15 centímetros de diámetro. Estas placas
integraban una coraza dérmica similar a la del Saltasaurus loricatus.
Powell describió en 1986 al saurópodo
titanosaurio Epachthosaurus sciuttoi basándose en una única vértebra
dorsal hallada en sedimentos de la Formación Barreal de la Estancia
Ocho Hermanos, Sierra de San Bernardo, provincia del Chubut. En
1975, Roberto Abel, entonces director del Museo Provincial de
Cipolletti “Carlos Ameghino”, descubrió unos restos de saurópodo que
Powell atribuyó al género Epachthosaurus. El hallazgo había sido
realizado en la localidad de Lago Pellegrini, provincia de Río
Negro, en sedimentos de la Formación Allen, depositados en el
Cretácico tardío. En un estudio posterior, Salgado interpretó que
este saurópodo rionegrino no correspondía al Epachthosaurus sciuttoi
y en 1996 describió a estos restos como pertenecientes a una nueva
especie, a la que denominó Pellegrinisaurus powelli.
Otros titanosaurios muy comunes en el
Cretácico tardío de Patagonia son el Titanosaurus araukanikus de
miembros esbeltos y el Neuquensaurus australis de características
similares al Saltasaurus loricatus. Ambas especies provienen de las
Provincias de Río Negro y Neuquén. Al igual que el resto de los titanosaurios, estas especies poseían vértebras de la cola con una
articulación posterior fuertemente convexa, lo que aparentemente
permitía una mayor movilidad de la cola.
El paleontólogo alemán Friederich von
Huene, de la Universidad de Tübingen, que fuera contratado por el
Museo de La Plata, efectuó un estudio de los dinosaurios del
Cretácico de la Argentina, que fue publicado en los Anales del
citado Museo de 1929. Esta obra constituye uno de los trabajos
clásicos sobre dinosaurios argentinos. Entre éstos se destacaba el
Antarctosaurus wichmannianus, descripto a partir de un esqueleto
incompleto colectado por el geólogo Wichmann, aproximadamente en
1916, en los sedimentos de la Formación Río Colorado, de edad
Conaciano –Cretácico tardío– del norte de Patagonia. Este gigantesco
saurópodo pertenece al grupo de los titanosaurios.
Otro titanosaurio gigante es el
Argyrosaurus superbus, descripto en 1893 por el paleontólogo inglés
Richard Lydekker a partir de una extremidad anterior completa
hallada por Carlos Ameghino en el río Chico de Chubut, próximo al
lago Musters. En 1978, una comisión de la Universidad Nacional de
Tucumán halló nuevos restos del Argyrosaurus superbus a orillas del
río Senguer, al sur de Colonia Sarmiento, provincia de Santa Cruz.
Estos materiales, junto con otros extraídos en el mismo lugar por el
Museo Argentino de Ciencias Naturales, fueron estudiados por Powell
en 1986. Los caracteres de las extremidades anteriores de este
titanosaurio sugieren que estaría relacionado con el Alamosaurus
sanjuanensis, un titanosáurido del Cretácico tardío de América del
Norte.
Los hallazgos de huevos y nidos de
dinosaurios cretácicos
Los dinosaurios fueron ovíparos y los
huevos a menudo se preservan como fósiles. En Argentina existen
varios yacimientos con huevos de dinosaurios de gran importancia,
aunque es muy difícil identificar al dinosaurio responsable de las
puestas.
Jaime Powell, de la Universidad Nacional
de Tucumán, fue quien dio el puntapié inicial en el estudio de los
huevos de dinosaurio en Argentina. Su área de trabajo se concentró
en el norte de la Patagonia, en un área ubicada aproximadamente al
sur de la localidad rionegrina de General Roca. Los nidos
descubiertos por Powell eran numerosos, con huevos de cáscaras
increíblemente gruesas y porosas. Powell entendió que el inusitado
grosor de las cáscaras constituía una protección segura contra los
depredadores.
Durante la reunión de comunicaciones
sobre paleobotánica y palinología, realizada en 1990 en la
Asociación Paleontológica Argentina, se dieron a conocer los
resultados de las investigaciones interdisciplinarias realizadas en
el Bajo de Santa Rosa, al norte de la localidad de Valcheta,
provincia de Río Negro. De este yacimiento del Cretácico tardío,
Bonaparte describió los restos de vertebrados fósiles y nidadas de
dinosaurios.
Teresa Manera de Bianco, de la
Universidad Nacional del Sur, dio a conocer en 1996 un nuevo
yacimiento con nidos y huevos de dinosaurios del Cretácico tardío
ubicado en la zona centro sur de la provincia de Río Negro, en un
lugar conocido localmente como Cerro Blanco. Las características de
estos huevos son similares a las de otros atribuidos tanto por Mones
como Powell a la familia de los titanosáuridos. La amplia difusión
de nidos en un mismo nivel estaría indicando una nidificación en
colonias, mientras que la ausencia de modificaciones en el sedimento
que rodea a las nidadas sugiere que los huevos eran enterrados en la
arena para su incubación. Además de los materiales mencionados, en
ese yacimiento se descubrieron impresiones de tallos y hojas de
plantas, troncos silicificados, restos de invertebrados, así como de
peces, tortugas, cocodrilos, dinosaurios saurópodos y terópodos.
En 1997, Calvo y Salgado, junto con
Silvia Engelland y Susana Heredia, de la Universidad Nacional del
Comahue, anunciaron el primer registro en Neuquén de cáscaras de
huevos de dinosaurios. Los afloramientos están localizados en la
ciudad de Neuquén y pertenecen a la formación Río Colorado, de edad
Coniaciano o Santoniano, con una antigüedad comprendida entre unos
88 a 83 millones de ańos. Los huevos probablemente eran esféricos,
con un diámetro estimado de 16 centímetros. De acuerdo a la
estructura de la cáscara, la semejanza con otros hallados en la
India y Francia, y a la presencia de restos de saurópodos en el
horizonte portador, los paleontólogos asignaron los huevos a estos
últimos.
Pero los hallazgos más espectaculares
serían realizados en 1998, cuando un grupo encabezado por Luis
Chiappe, del Museo de Historia Natural de Los Ángeles, y Rodolfo
Coria, anunció el descubrimiento de cientos de huevos conteniendo
embriones y restos de piel embrionaria. Esta es la primera vez en el
mundo que se encuentran restos de piel de embriones y la primera vez
en la Argentina que se descubren restos de embriones de dinosaurios.
żLos últimos dinosaurios carnívoros?
El primer terópodo registrado en América
del Sur es el Genyodectes serus, hallado a fines del siglo XIX en la
provincia de Chubut por Santiago Roth, del Museo de La Plata, y
descripto por Sir Arthur Smith Woodward, del Museo Británico.
Lamentablemente, tanto la posición estratigráfica como geográfica de
este fósil son desconocidas.
En los mismos sedimentos de la Formación
Barreal de la Estancia Ocho Hermanos en los que se halló al
saurópodo Epachthosaurus sciuttoi se descubrió el terópodo
Xenotarsosaurus bonapartei, descripto por R. Martínez, O. Giménez,
J. Rodríguez y G. Bochatey en 1986. El nombre genérico de este
dinosaurio significa lagarto con tarso extrańo y hace referencia a
la inusual conexión entre los huesos tarsales y la tibia.
El Miembro Portezuelo de la Formación
Río Neuquén aflora en la Sierra del Portezuelo, a unos 20 kilómetros
de Plaza Huincul, en la provincia del Neuquén. Estos sedimentos
corresponden a la edad Turoniana, con una antigüedad cercana a los
90 millones de ańos. En este yacimiento, sucesivas expediciones del
Museo Argentino de Ciencias Naturales “Bernardino Rivadavia”
dirigidas por Fernando Novas, descubrieron dos terópodos con
características avianas y una garra gigante, la más grande de las
conocidas, de un dinosaurio similar al Velociraptor, pero de un
tamańo mucho mayor (Megaraptor namunhuaiquii).
En 1997, Novas describió al Patagonykus
puertai, un dinosaurio con características avianas emparentado con
el Mononykus olecranus, de Mongolia. Este terópodo medía unos 2
metros de largo y poseía los brazos extremadamente cortos con dedos
atrofiados, excepto el pulgar, que estaba provisto de una garra
robusta. En el mismo ańo describió también al Unenlagia comahuensis,
otro dinosaurio con caracteres avianos también descubierto en la
Sierra del Portezuelo. La pelvis del Unenlagia es comparable a la de
la primitiva ave jurásica Archaeopteryx.
Una antigüedad algo menor tiene otro
dinosaurio aviano: el Alvarezsaurus calvoi, un terópodo del tamańo
de una gallina descubierto por Jorge Calvo en los terrenos de la
Universidad del Comahue, en Neuquén, y descripto por Bonaparte en
1991. Los sedimentos portadores de este fósil corresponden a la
Formación Río Colorado, de una edad probablemente Coniaciano, a la
que le corresponde una antigüedad de 88 a 86 millones de ańos. El
nombre genérico está dedicado al escritor e historiador neuquino
Gregorio Alvarez y el específico a su descubridor.
El Velocisaurus unicus también se
descubrió en los terrenos de la Universidad del Comahue. Este
terópodo, descripto por Bonaparte en 1991, está representado por una
extremidad posterior incompleta. El pie de este dinosaurio
representa una condición anatómica muy evolucionada, no registrada
en otros terópodos.
Otro pequeńo terópodo fue el Noasaurus
leali, descripto por Bonaparte y Powell en 1980. Este dinosaurio fue
descubierto en las capas de edad Maastrichtianas de la Formación
Lecho, en la Estancia El Brete, en el sur de Salta, con una
antigüedad de unos 70 millones de ańos. El Noasaurus leali
medía 1
metro de largo y poseía una garra recurvada para retener y matar a
sus presas. Este dinosaurio vivió junto con el saurópodo Saltasaurus
loricatus.
También procede de Salta el terópodo
Unquillosaurus ceibali, descripto por Powell en 1979 a partir de un
pubis izquierdo casi completo. Este pubis se halló en sedimentos de
la Formación Los Blanquitos.
Entre los últimos terópodos de gran
tamańo de la Argentina está el Abelisaurus comahuensis, que vivió
hace unos 70 millones de ańos. Este dinosaurio, que mediría unos 10
metros de largo, fue descubierto por el profesor Roberto Abel, ex
director del Museo Provincial de Cipolletti, en las cercanías del
lago Pellegrini, en la provincia de Río Negro, y descripto por
Bonaparte y Novas en 1985. El único resto que se conoce de este
dinosaurio es un cráneo incompleto que mide unos 85 centímetros de
largo. Otros dinosaurios afines al Abelisaurus comahuensis vivieron
en Madagascar, India y Europa.
Muchas de las características de las
aves, aún la presencia de plumas, son compartidas por ciertos
dinosaurios. Esto hace que muchos paleontólogos sostengan que las
aves son dinosaurios y que por lo tanto estos animales no se
extinguieron completamente a fines del Cretácico.
Agradecimientos
Los autores agradecen al doctor José
Bonaparte, del Museo Argentino de Ciencias Naturales “Bernardino
Rivadavia”, por la lectura crítica del primer manuscrito.
Bibliografía
José Bonaparte. “Dinosaurios del Jurásico de América
del Sur”. Investigación y Ciencia, Nş 63 (1981), páginas 110-121.
José Bonaparte. Los dinosaurios de la
Patagonia argentina. Buenos Aires. Asociación Herpelógica Argentina,
1985, Serie de Divulgación Nş 3, 31 páginas.
José Bonaparte. Dinosaurios de América
del Sur. Buenos Aires. Museo Argentino de Ciencias Naturales
“Bernardino Rivadavia”, 1996, 174 páginas.
José Bonaparte. El Triásico de San
Juan-La Rioja Argentina y sus dinosaurios. Buenos Aires. Museo
Argentino de Ciencias Naturales “Bernardino Rivadavia”, 1997, 190
páginas.
José Bonaparte. Los dinosaurios de la
Patagonia argentina. Buenos Aires. Museo Argentino de Ciencias
Naturales “Bernardino Rivadavia”, 1998, 46 páginas.
Jorge Orlando Calvo. Los dinosaurios del
período Cretácico de Patagonia. Neuquén. Universidad Nacional del
Comahue, 1995, 31 páginas.
Rodolfo Coria Y Leonardo Salgado.
“Dinosaurios carnívoros de Sudamérica”. Investigación y Ciencia, Nş
237 (1996), páginas 39-40.
Don
Lessem & Donald F. Glut. Dinosaur Encyclopedia. New York. The
Dinosaur Society’s, 1993, 533 páginas.
Alfredo Monetta.
Ischigualasto. Valle de Luna y dinosaurios. San Juan. Editorial
Fundación Universidad Nacional de San Juan, 1993, 66 páginas.
Fernando E. Novas. “Origen de los
dinosaurios”. Investigación y Ciencia, Nş 217 (1994), páginas 52-59.
Fernando E. Novas. Guía de los
dinosaurios de la Argentina. Buenos Aires. Edición del autor, 1996,
24 páginas.
Leonardo Salgado y Rodolfo Coria. Los
ornitópodos en Patagonia. Neuquén. Universidad Nacional del Comahue,
1993, 21 páginas.
|
Transgresiones y Regresiones Marinas en la Región Pampeana.
Geomorfología Regional.
Magnussen Saffer, Mariano
(2005). Transgresiones y regresiones
marinas en la Región Pampeana. Artículo divulgativo
publicado en PaleoWeb – Boletín Paleontológico. Ańo 3, 11: 32-35.
marianomagnussen@yahoo.com.ar
Aviso
Legal
Es muy
raro que aparezca un fósil de origen marino en una zona terrestre. La
explicación, proviene de los diferentes estados naturales que se han
suscitado en el transcurso de los milenios.
Localmente, se denominan transgresiones marinas o ingresiones marinas a
los avances de la línea de costa sobre un espacio continental, y
regresiones marinas al fenómeno inverso.
La
orilla del mar es "fluctuante", es decir, en la época de los glaciares
el borde costero de la región pampeana se encontraba a 200 kilómetros
mar adentro, sobre el borde de la actual plataforma marítima, ya que el
agua se depositaba en aquellos fenómenos naturales, llamados glaciares,
lo cual impedía una entrada de mayor altitud.
|
 |
De la
misma manera, un interglaciar (el momento opuesto) hubo más agua, y hubo
momentos en que el mar subió hasta seis metros sobre el nivel actual, de
modo que lo que hoy es tierra firme, en otro tiempo se hallaba por
debajo del nivel de mar.
Durante
el Mioceno una gran parte de la Argentina estuvo cubierta por un mar
conocido ampliamente en la literatura geológica como "mar entrerriense o
paranense", del que se conocen facies marinas en el este y facies
marginales y lacustres en el oeste y nororeste del país. Los espesores
más potentes corresponden a los depósitos del subsuelo del ámbito Chaco-paranense,
La
característica más notoria de todos estos depósitos lo constituye la
abundante y diversa fauna de moluscos restringida a las facies marinas
de las formaciones Paraná y Puerto Madryn y a las del subsuelo de la
provincia de Buenos Aires.
|
Las
primeras menciones sobre la existencia este mar son de índole
paleontológica y datan desde mediados del siglo diecinueve. Fueron
efectuadas por d’Orbigny (1842), quien halló moluscos marinos en los
alrededores de la ciudad de Paraná y en la desembocadura del río Negro.
Unos ańos más tarde, Darwin (1846) encontró la misma fauna en la
península Valdés, quedando así establecida la presencia de estos
depósitos en la Patagonia septentrional y en la provincia de Entre Ríos,
los que constituyeron, por otra parte, las primeras citas del Terciario
marino en la Argentina.
|
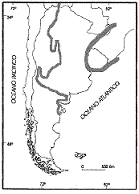 |
En la
región mesopotámica, los depósitos miocenos corresponden a la Formación
Paraná Yrigoyen (1969) y se restringen al borde oriental de la misma,
aflorando a lo largo del río Paraná entre Diamante y La Paz. A
diferencia de la Formación Puerto Madryn, esta unidad aflora en forma
discontinua y se caracteriza por una marcada variación facial. De
acuerdo con Aceńolaza (1976), la sección aflorante de la Formación
Paraná no supera los 30 metros de espesor y está constituida por
arcillas, arcillitas limolíticas verdosas, arenas, areniscas limolíticas
amarillentas y coquinas con matriz arenosa o calcárea.
|
Con
respecto a los ambientes de depositación de las Formaciones Puerto
Madryn y Paraná en sus áreas tipo, los análisis paleontológicos y
paleoambientales realizados seńalan la presencia de facies marinas
someras para los términos de la transgresión, que se extienden desde la
Patagonia hasta la provincia de Entre Ríos, a lo largo de una franja de
rumbo norte-sur, de aproximadamente 1100 kilómetros de longitud.
La
Formación Paraná habría sido depositada en un ambiente marino somero con
influencias deltaicas (Iriondo, 1973). Por otra parte, Aceńolaza y
Aceńolaza (2000) determinaron que esta unidad correspondería a una
secuencia transgresiva depositada desde ambientes aéreos o subaéreos a
submareales, representados estos últimos por barreras arrecifales.
|
 |
Con
respecto a la Formación Puerto Madryn aflorante en los alrededores de la
ciudad de Puerto Madryn y la península Valdés, se determino que se trata
en términos generales de una secuencia regresiva con facies depositadas
en la plataforma por debajo de la base del tren de olas en ambientes de
baja energía, y de facies más someras depositadas en un ambiente con
dominio alternativo de mareas y tormentas (facies de lenguas arenosas
submareales, de canales de mareas, coquinas tempestíticas y planicies de
mareas) y paleosuelos. |
A
conclusiones similares arribaron Zucol y Brea (2000), quienes
determinaron condiciones tropicales a subtropicales húmedas para el
palmar desarrollado en el ambiente marino costero donde se habrían
depositado las sedimentitas de la Formación Paraná.
La
transgresión marina llamada Mar Paranense habría comenzado hace unos 15
y 14 Millones de ańos, y habría concluido hace 7
y 6 Millones de ańos abarcando desde el mioceno medio al tardío
(algunos autores sostienen hasta comienzos del plioceno).
La
ingresión de este mar se dio por el río de la Plata y la cuenca del río
Salado (Provincia de Buenos Aires).
Era un
mar somero, de aguas con temperaturas levemente superiores a las
actuales y de una gran extensión areal, abarcando la casi totalidad de
la llanura chaco - pampeana y extendiéndose hasta el sur de Brasil,
Bolivia y Paraguay. El flanco oeste tenía sus costas en las cercanías de
las sierras Pampeanas, mientras que la costa oeste llegaba hasta el
actual río Uruguay. El flanco norte llegaba hasta la región del
Pantanal, en la parte superior del río Paraguay.
La
concentración salina habría ido en descenso hacia el interior del
continente, sobre todo por los aportes de agua dulce de los numerosos
ríos que desembocaban en el citado mar.
Los
movimientos de levantamiento andino en el mioceno medio habrían sido
causales del hundimiento de la llanura chaco pampeana, permitiendo así
la ingresión de aguas del Océano Atlántico al continente, mientras que
los movimientos de elevación de las Sierras Pampeanas (que ya existían
desde tiempos inmemoriales, pero por acción del levantamiento andino
sufrieron nuevos elevamientos y fracturación de sus bloques formantes) a
finales del mioceno habrían contribuido a la regresión de este mar.
 |
Durante
el Cuaternario también hubo algunas transgresiones marinas (pero no tan
importantes como la anterior) asociadas sobre todo a los periodos
interglaciares, donde el agua de deshielo de los casquetes polares
aumentaba los niveles de los océanos. Una de las más recientes fue la
denominada Mar Querandí, que ocurrió hace 7500 a 4000 ańos atrás,
inundando el estuario de la Plata (que fue por donde ingresó) y llegando
por el río Paraná hasta la altura de la ciudad de Diamante, Entre Ríos.
|
Durante
el Cuaternario también hubo algunas transgresiones marinas (pero no tan
importantes como la anterior) asociadas sobre todo a los periodos
interglaciares, donde el agua de deshielo de los casquetes polares
aumentaba los niveles de los océanos. Una de las más recientes fue la
denominada Mar Querandí, que ocurrió hace 7500 a 4000 ańos atrás,
inundando el estuario de la Plata (que fue por donde ingresó) y llegando
por el río Paraná hasta la altura de la ciudad de Diamante, Entre Ríos.
Cabe
destacar que en la historia hubo varias ingresiones (o entradas de mar)
registradas. Una de ellas data de 120.000 ańos y la segunda es la que
dio origen a la laguna de Mar Chiquita y los sedimentos de Punta
Hermengo hace 6.000 ańos (Formación Querandi).
|
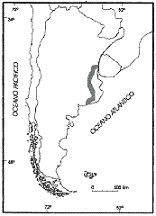 |
En la
zona costera de Buenos Aires se reconocen varios niveles
correspondientes a otras tantas probables oscilaciones del nivel
marítimo. A lo largo de la costa atlántica del NE Argentino, el nivel
mas antiguo se encuentra en el interior de la Formación Ensenada, al
cual Ameghino (1889) llamo Intersenadense. Esta
transgresion, que se desarrolla a lo largo de la desembocadura del Rió
de La Plata. Otras tres pulsaciones marinas sucesivas han sido
descriptas en esta zona costera: Belgranense, Querandinense y Platense
marino.
En la
zona de Miramar, se conocen depósitos referibles a la ingresion
holocenita, en particular, en la serie expuesta en la desembocadura del
Arroyo Las Brusquitas, y en facies salobres en Punta Hermengo. Ambos
poseen una antigüedad de 6000 ańos.
|
La
serie de Las Brusquitas, situada sobre la costa atlántica Argentina, en
la desembocadura del arroyo homónimo, pertenece a un ambiente marino –
salobre en el cual se suceden indicios de oscilaciones del nivel del
mar, bajo forma de depósitos con fósiles de ambientes claramente marino
costero, intercalado con depósitos fluviales y salobres.
 |
Se
pueden encontrar en estos sedimentos, restos óseos de distintos
vertebrados, pero escasos. Se caracteriza por la gran variedad de
moluscos, como Biomphalaria, Littoridina parchappei y
Littoridina
australis entre otras, abundantes en el perfil y representativas de todo
los ambientes, dulceacuicola, salobre y mas ampliamente marino.
<<<<Cordón de conchillas del holoceno de "Las Brusquitas". |
Hace aproximadamente 8000 ańos y tras una importante
desglaciación, el mar (formación Querandi) llegó hasta lo que ahora es la Ruta Nacional Nş 2
y desde el río Salado hasta San Pedro (Provincia de Buenos Aires). Cuando las aguas se retiraron
(hace unos 4000 ańos), el terreno adquirió una fisonomía heterogénea.
Los sedimentos salinos formaron mantos geológicos capaces de alterar el
sabor y las características de las aguas en las zonas deprimidas o bańados,
esto justifica la aparición reiterada de la palabra "salado" en
nombre de ríos, arroyos y lagunas.
Bibliografía Sugerida.
Aceńolaza,
F.G. 1976. Consideraciones bioestratigráficas sobre el Terciario marino
de Paraná y alrededores. Acta Geológica Lilloana 13 (2): 91-107.
Tucumán.
Aceńolaza,
F.G. 2000. La Formación Paraná (Mioceno medio): estratigrafía,
distribución regional y unidades equivalentes. En: Aceńolaza F. G. y
Herbst, R. (eds.), El Neógeno de Argentina. Correlación Geológica 14:
9-27.
Aguirre, M.
L. 1990. Asociaciones de moluscos bentónicos marinos del cuaternario
tardío en el noroeste bonaerense. Ameghiniana 27: 161-177.
Alberd M. T.
i, Leona G. y Tonni E. P. (editores). Evolución biológica y climática de
la región pampeana durante los últimos cinco millones de ańos. Un ensayo
de correlación con el Mediterráneo occidental. Madrid, Museo Nacional de
Ciencias de Madrid, Consejo Superior de Investigaciones Científicas,
1995, Monografías 12.
Bravard, A.
1858. Monografía de los terrenos marinos terciarios de las cercanías del
Paraná. Imprenta del Registro Oficial. 107 pp. Paraná. (Reimpresión
Imprenta del Congreso de la Nación, 1995).
Cozzuol, M.A.
1993. Mamíferos acuáticos del Mioceno medio y tardío de Argentina.
Sistemática, evolución y biogeografía. Tesis Doctoral, Facultad de
Ciencias Naturales y Museo, UNLP. 178 pp. Inédita.
Francesco
C.G. y Zarate, M.A.1999. Análisis tafonomico de Littoridina Souleyet,
1852 (Gastropoda: Hydrobiidae) en perfiles holocenos del rio Quequen
grande (Prov. Buenos Aires): significativo paleobiologico y
paleoambental. Ameghiniana 36 (3): 297-310 Bs.As.
Fidalgo, F.;
Tonni, E.P. 1983. Geología y Paleontología de los sedimentos encauzados
del Pleistoceno tardío y Holoceno en Punta Hermengo y Arroyo Las
Brusquitas (Partido de General Alvarado y General Pueyrredón, Provincia
de Buenos Aires). Ameghiniana, Vol. 20, Nos. 3-4, p. 281-296.
Firinati,
E.A. y Aliotta, S.A. 1995. Análisis tafonomico de conchillas en cordones
holocenos, Bahía Blanca, Argentina. 4 Jorn. Geológicas y geofísicas
bonaerenses (Junin) Actas 1: 89 – 97.
Herbst, R. y
Zabert, L.L. 1987. Microfauna de la Formación Paraná (Mioceno superior)
de la cuenca Chaco-Paranense (Argentina). Facena 7: 165-206. Corrientes.
Tonni E.P. y
Cione, A.L. Los mamíferos y el clima en el Pleistoceno y Holoceno en la
provincia de Buenos Aires.- Jornadas de Arqueología e Interdisciplinas,
PREP, CONICET, pp. 127-142; Climas en el Cuaternario de la Región
Pampeana y cambio global. II Seminario sobre las Geociencias y el cambio
global, Asoc. Geol. Arg., Serie D, Publ. esp. N° 2:33-35; 1995.
El largo viaje al
Plata del Diplodocus carnegii.
Por
Irina Podgorny de la UNLP-CONICET y
Tobías Plöger de la
Universität Köln.
Fragmento del articulo publicado en la Revista de la Fundación
Ciencia Hoy.
Volumen 9 - Nş 51 MARZO-ABRIL 1999.
Primera imagen original del articulo. Las siguientes son del archivo
de PaleoArgentina Web.
Aviso
Legal
Desde 1912 se exhibe en una sala del
Museo de La Plata el regalo que el multimillonario norteamericano
Andrew Carnegie le hizo al entonces presidente Roque Sáenz Peńa. Se
trata de un calco de la reconstrucción del enorme dinosaurio
Diplodocus carnegii,
y cuyo nombre es un homenaje a Carnegie por su mecenazgo al Museo de
Pittsburgh y a la paleontología norteamericana. La réplica es
similar a las que existen en varios museos de historia natural de
Europa, pero es la única en América Latina. La donación puede
entenderse como parte de la misión de paz que las instituciones
creadas y mantenidas por Carnegie pretendían impulsar en los ańos
anteriores a la Primera Guerra Mundial. El reptil fósil
norteamericano llegaba a la Argentina para compartir las salas con
los grandes mamiferos autóctonos que expresaban la riqueza
fosilífera de esta parte del continente; en el Norte, en cambio, los
hallazgos de las expediciones paleontológicas norteamericanas a las
montańas Rocallosas, rivalizaban con los dinosaurios de Inglaterra y
Europa continental.
La categoría de los Dinosauriae fue creada en 1841 por Richard
Owen, el especialista inglés en anatomía comparada. Comprendía a
reptiles de enorme tamańo, cuello y cola largos, y cráneo pequeńo en
comparación con el resto del cuerpo. El orden se conformaba con
especimenes de formas extremadamente variadas que vivieron en el
Mesozoico (245 a 66 millones de ańos). Su distribución mundial quedó
demostrada ya a principios del siglo XX, por los hallazgos en toda
América, Madagascar, China, India, Africa oriental y Europa.
Recordemos que los primeros restos fueron descubiertos a comienzos
del siglo pasado en las capas del Jurásico (208 a 114 millones de
ańos) y del Wealden (144 a 138 millones de ańos) ingleses y que las
primeras descripciones las realizaron Buckland en 1824 y Mantell en
1825. A la gran tarea sistemática de Richard Owen, deben sumarse las
de Thomas Henry Huxley, John Phillips, Edward Cope y Othniel Marsh.
Fue este último quien los dividió en tres subórdenes: Theropoda,
Saurapoda y Predentata; mientras que H.G. Seeley los agrupó en
Saurischia y Ornithischia. En el suborden de los Sauropoda -el grupo
menos especializado dentro de los Saurischia-, se constituyó la
familia de los Diplodocidae, con el género Diplodocus,
a partir de los restos hallados en el Jurásico Superior (163 a 144
millones de ańos) de la Formación Morrison en Colorado y Wyoming (EE.UU.).
El Diplodocus era el más especializado de los
saurópodos, lo que se hacia patente por la forma elongada de las
regiones caudales y cervicales, lo escueto de la porción
dorso-lumbar, la estructura compleja de las vértebras, la marcada
reducción del número y del tamańo de los dientes, y por la
naturaleza reducida de los dígitos III, IV y V en el pie.
Othniel Marsh, profesor de
la Universidad de Yale (EE.UU.), fue quien inventó el nombre
de Diplodocus.

DIPLODOCUS CARNEGII.
REPRODUCCIÓN DE UN EJEMPLAR PROCEDENTE DEL JURÁSICO
SUPERIOR,
WYOMING (ESTADOS UNIDOS). MUSEO DE HISTORIA NATURAL, LONDRES |
El, gracias a la fortuna que le
había legado su tío George Peabody, pudo costear sucesivas
expediciones a las Rocallosas para buscar los preciados
restos de dinosaurios. También creó en 1878 el género
Diplodocus a partir de las vértebras de la cola y
fragmentos de las patas que halló su equipo en los estratos
del Jurásico del estado de Colorado (EE.UU.).
Marsh compuso el nombre a partir
de dos palabras griegas: diploös, doble, y dokos, viga o
bao; se inspiró en los "chevrones" de las vértebras de la
parte baja de la cola del reptil, que se presentaban en
pares a semejanza de la estructura de las vigas de un navío.
La forma y la estructura del animal se dio por resuelta con
los hallazgos sucesivos de una pelvis y de gran parte de la
cola, descriptas por Henry Fairfield Osborn, paleontólogo
del Museo Americano de Historia Natural de Nueva York y
heredero de una familia millonaria. |
Las copias no fueron recibidas de la misma manera en todos lados. En
Alemania, particularmente, se desató un intenso debate acerca del diseńo
del Diplodocus. Para algunos paleontólogos alemanes, los norteamericanos
habían caído en el error de presentar las patas del reptil a la manera de
los paquidermos, cuando hubiese sido más apropiado concebirlas como las de
los cocodrilos. También fue cuestionada la forma del cuello y de los pies,
pero finalmente triunfó la interpretación consagrada por la réplica hecha
en Pittsburgh. Este debate ocupó varias sesiones de la "Sociedad de amigos
de las ciencias naturales de Berlin", además de muchas páginas en las
revistas científicas alemanas y norteamericanas, que prolongaba, de alguna
manera, la disputa entre Owen y Huxley sobre el lugar de los dinosaurios
en el orden natural. Por otro lado, los mismos críticos seńalaban que los
científicos franceses sólo se habían preocupado por la estética de la
instalación, descuidando el problema más polémico: la ligereza de los
norteamericanos en la interpretación de la forma arquetípica de los
dinosaurios.
El Diplodocus en el Plata.
Si se tiene en cuenta que la tarea filantrópica y científica de
Carnegie estaba orientada más bien hacia Europa, resulta extrańo la
donación del Diplodocus a un museo sudamericano. Por otra parte, además de
los museos y del Instituto Carnegie en Pittsburgh, los fondos del
millonario se destinaban a la investigación científica, entre las que se
cuentan las excavaciones de sitios arqueológicos en México y
Centroamérica, y a la creación en los Estados Unidos, Europa y África de
bibliotecas para la educación de nińos con pocos recursos. La llegada del
Diplodocus a las orillas del Plata fue un acto excepcional, no solo para
los fondos de Carnegie, sino incluso para las mismas instituciones
científicas argentinas.
Desde 1906 el Museo de La Plata formaba parte de la nueva universidad
nacional fundada por Joaquín V. González, y se había establecido contando
con las colecciones con que en 1877 Francisco Moreno había creado el Museo
Antropológico de la Provincia de Buenos Aires. El volumen de las mismas no
era suficiente para completar las salas del edificio monumental que se
levantó en 1884 en la nueva capital provincial: las cartas de Moreno
hablan a las claras de la premura con que tuvo que reunir los materiales
para llenar esos enormes espacios que hacían parecer pequeńos hasta los
magníficos gliptodontes pampeanos. Aún en la primera década de este siglo
había salas casi vacias, razón por la cual en 1911 Lehmann-Nitsche, jefe
de la Sección de Antropología, inició las gestiones ante la embajada
argentina en Washington para recibir una reproducción del Diplodocus
carnegii. Ya antes se había dirigido a Mr. Robert Woods Bliss, encargado
de negocios de los Estados Unidos en la Argentina, quien en la respuesta
oficial del 10 de enero de 1911, comenta: "Acuso recibo de su carta de
fecha 5 en relación a su deseo de que el Sr. Andrew Carnegie regale al
Museo de La Plata un modelo de yeso del Diplodocus carnegii, similar a las
reproducciones que el Sr. Carnegie ya ha donado a varios museos europeos.
Ud. manifiesta que al presente hay en el museo una sala del tamańo
adecuado para albergar la reproducción que solicita, y ańade que no sería
difícil obtener dicha reproducción por vía diplomática ya que aún existen
los moldes originales". En la carta se destaca que el pedido no lo inician
las autoridades del museo ni de la universidad, sino el jefe de la Sección
de Antropología, que no era el responsable de la sala que se ponía a
disposición. La embajada norteamericana no aceptó oficialmente el trámite,
dado que se trataba de interceder ante un particular: "Esta legación, sin
embargo, no puede presentar una solicitud a un particular pidiendo un
regalo para una institución del país donde se encuentra la legación". De
todos modos, Woods Bliss le aconsejó a Lehmann-Nitsche cuáles podrían ser
los canales para lograr la donación. Las gestiones tuvieron éxito y en
septiembre de 1912 llegaban al país el director del Museo de Pittsburgh,
William Holland, y su preparador en jefe, Arthur Coggeshall, para ocuparse
de la instalación del Diplodocus. Ambos habían montado el modelo en los
museos europeos y acumulaban práctica y experiencia -además de
accidentes-, como para relatar innumerabIes anécdotas a quien quisiera
escucharlos. Los norteamericanos fueron alojados en una casa del
Observatorio Astronómico, a corta distancia del museo. Como en todos los
casos anteriores, el Diplodocus tenía el carácter de regalo al primer
mandatario del país.

El técnico Mariano Magnussen Saffer, junto al
ejemplar de Diplodocus
carnegii exhibido en el Museo de La Plata en la actualidad. (2018) |
La instalación duró aproximadamente un mes y pasó absolutamente
inadvertida para los medios periodisticos; sólo Caras y Caretas envió un
corresponsal a La Plata para conversar con el Dr. Holland, y publicó un
artículo profusamente ilustrado con una nota autografiada donde el
norteamericano expresaba su deseo de "reencarnar en este suelo". La
ausencia de noticias sobre la llegada a La Plata del colosal dinosaurio no
puede adjudicarse a la falta de interés por las ciencias naturales, tanto
los diarios platenses como los porteńos publicaban las novedades que
ocurrían en los jardines zoológicos y los hallazgos de fósiles en
distintas regiones del país. Pero sí se ocupaban en extenso de los eventos
sociales que acompańaban al ceremonial de la ciencia. Aquí a diferencia de
las inauguraciones europeas, no hubo festejos en el museo sino que se
organizaron sucesivos banquetes en honor al Dr. Holland.
|
En ninguno de ellos participó el presidente de la Nación quien, no
obstante, recibió al paleontólogo en su despacho de la casa de gobierno
el 15 de octubre de 1912.
La visita fue casi ignorada por los diarios de Buenos Aires, que
sólo la consignaban como otorgada a "Juan Holland" en la escueta lista de
las cinco audiencias presidenciales del día. Esta circunstancia contrasta
con lo que asienta William Holland en su diario, donde pondera la cultura
y el manejo del inglés de Sáenz Peńa. Recordemos que de regreso a su país,
Holland publicó las impresiones del viaje en el libro To the River Plate
and back, donde se extendía sobre las contradicciones que había observado
en la sociedad argentina. A modo de testimonio documental, se incluían en
el volumen fotos del Teatro Colón y del Jockey Club, junto con las
viviendas de los sectores más pobres de la ciudad de Buenos Aires.
El 24 de octubre de 1912, el Consejo Superior de la Universidad
Nacional de La Plata organizó en el salón imperial del Jockey Club el
banquete de despedida al "ilustre huésped", al que fueron invitados
funcionarios nacionales y provinciales, los embajadores de Estados Unidos,
Chile y el Reino Unido, profesores universitarios y directores de museos.
En el brindis que honraba al enviado de Carnegie, Joaquín V. González se
explayó sobre el papel que le cabía a la ciencia en el fortalecimiento de
la paz internacional: "Tengo entendido que si las convenciones políticas
fundadas en la sola voluntad e intereses transitorios de las partes pueden
crear períodos más o menos duraderos de paz, sólo la ciencia puede
hacerlos estables y permanentes, porque sólo ella es capaz de crear una
armonía perfecta de todos los hombres, porque los hace conocerse los unos
a los otros -lo que es amarse-, y los hace comprender en qué y por qué son
todos iguales: lo que quiere decir suprimir la causa generadora de todo
conflicto humano y de toda perturbadora ambición. El Diplodocus carnegie o
Pacificus se halla ya instalado en el Museo de La Plata, entre una pléyade
colosal de sus contemporáneos patagónicos para dar fe en todo tiempo
venidero, de una remotísima época acerca de la cual, si no me atreviera a
afirmar que reinase siempre la paz, aun fundada en el equilibrio de las
fuerzas físicas, no vacilaría en decir que por lo menos todos los seres de
la escala superior, incluso el hombre, podían sin rubor alguno llamarse
grandes; mientras que los otros, los de la época actual, con toda la
ciencia, las religiones y las filosofías acumuladas, apenas podemos
comprendernos, igualarnos artificialmente, y vivir en paz unos al lado de
otros, en nuestra péqueńez reconocida". Con este brindis, González parecía
querer celebrar los vínculos de solidaridad establecidos por la práctica
científica internacional y la creación en el museo de un monumento a la
convivencia pacífica entre los pueblos americanos. De esta manera, según
su retórica científico-espiritualista, la réplica industrial del viejo
esqueleto se incorporaba a un paisaje idealizado de virtudes morales, para
instalar en las salas del museo un símbolo de la paz que se legaba al
futuro.
|

Mariano Magnussen Saffer
junto a dos fémures de Antarctosaurus
en el Museo de La Plata. (archivo) |
A los pocos ańos, la permanencia pacífica del
Diplodocus en las salas de paleontología del museo fue perturbada. En 1920
fueron desenterrados en Neuquén los restos de un dinosaurio que pasó a
denominarse Antarctosaurus. Los huesos de las patas medían más de dos
metros, con lo que superaba en tamańo al Diplodocus y al Gigantosaurio
africano: el "dinosaurio argentino" pasaba a ser el más grande de todos
los tiempos y de toda la Tierra. La Sección de Paleontología del Museo de
La Plata aprovechó la estructura del dinosaurio norteamericano para montar
al lado de sus patas traseras los dos fémures del Antarctosaurus, de manera
tal que la imaginación del público pudiera concebir cuán grande debió
haber sido el reptil local en comparación con su símil estadounidense.
Este acto de orgullo nacional por poseer el dinosaurio más grande, hacía
evidente la imposibilidad de reconstruir en la Argentina un modelo
completo, no sólo por la falta de recursos técnicos, sino fundamentalmente
por la desprotección económica en que se encontraba la investigación
científica. La reproducción en serie del Diplodocus carnegii
fue posible gracias a un proceso de institucionalización de las
ciencias que se fundaba en la estrecha alianza entre los
organismos de investigación y el capital privado.
|
En este sentido, la exaltación de la grandeza natural
estadounidense era sólo una excusa para propagar por el mundo entero la
gloria industrial del sueńo americano. Las patas del Antarctosaurus, en
cambio, daban al Diplodocus de La Plata la apariencia de un monstruo de
seis patas: el dinosaurio argentino quedaba anclado en los sueńos
retóricos sobre la grandeza del pasado.
La Primera Guerra Mundial acabó con los sueńos de paz y amistad
internacional promovidos por la ciencia y, en consecuencia, en la
posguerra se afianzaron definitivamente los principios de clausura y
rivalidad entre las tradiciones científicas de cada país. Andrew Carnegie
murió en 1919, cuando ya era evidente el fracaso de la misión de paz de su
Diplodocus.
LECTURAS SUGERIDAS
DESMOND, ADRIAN, 1979, "Designing the Dinosaur: Richard
Owen's Response to Robert Edmond Grant", Isis, 70: 224-234.
DESMOND, ADRIAN, 1982, Archetypes and Ancestors. Paleontology
in Victorian London 7850-7875, The Chicago University Press,
Chicago.
KOHLER, ROBERT E., 1991, Partners in Science. Foundations and
Natural Scientist 1900-1945, The Chicago University Press,
Chicago.
RUDWICK, MARTIN J.S., 1972, The meaning of Fossils: Episodes
in the History of Paleontology, MacDonald. (Hay traducción
espańola), Londres.
RUDWICK, MARTIN J.S., 1992, Scenes from the deep time, The
Chicago University Press, Chicago
Diario de un Caracol: cómo era el clima en el
pasado.
Sobre
Investigación. Claudio De Francesco. Investigador Adjunto.
Instituto de Geología de Costas y del Cuaternario (Universidad
Nacional de Mar del Plata). Fuente: Por Elena Aga (Mar del
Plata)
prensa@conicet.gov.ar.
Imágenes ilustrativas de nuestro Archivo.
Aviso
Legal
El
investigador del Conicet Claudio De Francesco afirma que los
caracoles de agua dulce que habitan en el centro-oeste de
Argentina son buenos indicadores para inferir las condiciones
ambientales del pasado, con registros que abarcan más de 35.000
ańos.
Los
caracoles que habitan los cuerpos de agua continentales como
ríos, arroyos y lagunas están comúnmente representados como
fósiles en la mayoría de los afloramientos de edad cuaternaria
de la Argentina, lo cual nos permite reconstruir cómo fue el
clima en el pasado. żDe qué manera? Conociendo el ambiente en el
que vive cada especie de caracol en la actualidad.
 |
Desde 2007 se ha comenzado a estudiar la relación entre
los caracoles y las principales variables ambientales
que pueden condicionar su distribución, a lo largo de
una gradiente de cuerpos de agua tales como ríos,
lagunas, arroyos, embalses y canales. El estudio se está
desarrollando en la provincia de Mendoza y el oeste de
San Luis.
Los
resultados indican que los caracoles en general
prefieren cuerpos de agua muy superficiales, con
abundante vegetación acuática, de baja salinidad, de
fondos fangosos y baja o nula velocidad de corriente. |
Una
excepción a la regla es el caracol Heleobia parchappii, que
tiene la capacidad de tolerar salinidades altas, incluso de
valores similares a las del mar. Por esta razón es muy abundante
y usualmente la única especie presente en las numerosas lagunas
salobres presentes en las provincias de La Pampa, San Luis,
Mendoza y Buenos Aires, constituyendo un buen indicador de este
tipo de ambientes.
De acuerdo a
los estudios de De Francesco se pueden reconocer tres
agrupaciones principales de caracoles: la parchappii,
restringido en aguas salobres; el Lymnaea viator, Biomphalaria
peregrina y Physa acuta, característico de ambientes lacustres o
de muy baja velocidad de corriente (arroyos pequeńos de bajo
caudal), y el Heleobia hatcheri, Heleobia cf. kuesteri, Chilina
mendozana y el bivalvo Pisidium chiquitanum, característico de
ambientes con mayor velocidad de corriente, como ríos y arroyos.
 |
Los
tres grupos de caracoles definidos a partir de los
estudios actuales se reconocen también en los depósitos
fosilíferos de las provincias de Mendoza y San Luis (con
la excepción de las especies P. acuta y H. hatcheri),
permitiendo aportar abundante y valiosa información para
bosquejar un modelo de evolución paleoambiental para el
Pleistoceno tardío (35.000-10.000 ańos) y Holoceno
(últimos 10.000 ańos) de la región. Así, el registro de
caracoles más antiguo está dado para los 35.000-31.000
ańos a.p., representado por las especies L. viator y B.
peregrina (segundo grupo del modelo actualista explicado
arriba) que indica un momento de energía relativamente
baja. |
Se trataría
de pequeńos encharcamientos, probablemente originados en
meandros abandonados de ríos, que habrían estado sujetos a
sumersiones ocasionales dentro de un régimen semi-temporario.
Este momento habría sido más cálido y húmedo que aquellos
registrados para el resto del Pleistoceno.
Entre los
30.000 y los 20.000 ańos a.p. no se registran caracoles, lo que
podría vincularse a un incremento en la aridez regional y
descenso de la temperatura. A partir de los 20.000 ańos a.p. se
registra por primera vez C. parchappii (indicador de aguas con
velocidades de corriente moderadas a altas) en Salinas del
Bebedero.
Con
posterioridad, este caracol se registra de manera casi continua
hasta finales del Pleistoceno, lo que respondería a un
incremento en la circulación efectiva de agua en toda la cuenca.
Los ríos habrían desarrollado un importante caudal para este
momento que, sumado al escaso desarrollo de la vegetación debido
a las condiciones de aridez imperantes, habrían resultado en
condiciones de alta energía. Esta situación se habría mantenido
hasta comienzos del Holoceno.
 |
A
partir de los 10.000 ańos a.p. se registran además B.
peregrina y H. parchappii tanto en San Luis como en
Mendoza, lo que se interpreta como momentos de
alternancia entre condiciones de alta circulación de
agua en la cuenca y episodios de encharcamientos
(probablemente en la llanura de inundación del cauce
principal), esto último indicativo de una disminución en
la energía relativa del ambiente.
Entre los 8000 y 4000 ańos a.p. (Holoceno medio) no se
registran caracoles, lo que podría deberse nuevamente a
la existencia de condiciones áridas. |
Finalmente,
durante el Holoceno tardío (últimos 4000 ańos) se produce un
mejoramiento ambiental evidenciado por la mayor abundancia y
diversidad de caracoles dulceacuícolas. En general, el panorama
paleoambiental para este momento es de poca circulación de agua
efectiva, dando origen a encharcamientos o pequeńas lagunas, muy
vegetadas, donde las especies hallarían un hábitat adecuado.
Este modelo
de inferencia paleoambiental está actualmente siendo ajustado a
través de la incorporación de otros aspectos preservados en las
conchas de los caracoles que pueden dar idea de cómo fue el
ambiente, como son los procesos de alteración sufridos desde la
muerte hasta la fosilización (procesos tafonómicos) y la
composición mineralógica, principalmente en algunos elementos
químicos (carbono y oxígeno) que están directamente relacionados
al clima del momento.
Fuente:
Conicet.
El origen de las flores.
Por Ari Iglesias y Mauro
Passalia,
Paleontólogos del INIBIOMA (CONICET-UNCOMA).
Originalmente publicado en ANB, y adaptado para este medio.
ari_iglesias@yahoo.com.ar Aviso
Legal
Hoy en día,
al llegar la primavera podemos ver flores de muy variadas
formas y colores. El resultado de la fecundación de las
flores da lugar a llamativos frutos conteniendo las
semillas, que al germinar darán origen a una nueva planta,
completando así el ciclo reproductivo del gran grupo de
plantas con flor que denominamos “angiospermas”.
Actualmente, las angiospermas, presentan una notable
diversidad de hábitos de crecimiento y dominan prácticamente
todas las comunidades vegetales (ˇhay angiospermas incluso
en la Antártida!). Agrupadas en 400 familias, se estima que
este grupo de plantas está compuesto por de 300.000
especies, lo cual excede la diversidad específica conjunta
de otros grupos de plantas terrestres.
No
obstante, este mundo florido tal como lo conocemos
actualmente, no siempre ha sido así. A lo largo de la
historia de la Tierra, las comunidades vegetales estuvieron
dominadas por diferentes grupos de plantas que le daban un
aspecto muy diferente al que hoy conocemos. De hecho, desde
que las primeras plantas colonizaron los ambientes
terrestres hace unos 450 millones de ańos, debieron pasar
300 millones de ańos más de evolución para que aparezcan las
primeras flores.
Las
angiospermas se diferencian de otras plantas terrestres
-como por ejemplo las coníferas, los helechos y los musgos-
por desarrollar flores. Sin embargo hay otras diferencias
vinculadas a su reproducción, al desarrollo de tejidos para
el transporte de agua y azúcares en la planta, como así
también en la capacidad de adaptarse a distintos hábitos de
vida.
żQué es una flor ?
Las flores
son los órganos reproductivos de las angiospermas.
Básicamente consisten en estructuras portadoras de uno o
varios ciclos de brácteas (pétalos y sépalos) y órganos
fértiles masculinos (el androceo, consistente de estambres
productores de polen) y femeninos (el gineceo, que envuelve
y protege a los óvulos que luego generarán los frutos y las
semillas). La mayoría de las angiospermas tienen flores
hermafroditas (con los dos sexos), pero hay especies que
desarrollan sus órganos masculinos y femeninos en flores
separadas.
|
 |
La
evidencia inequívoca más antigua de una planta con
flor es del período Cretácico Inferior (hace 140
millones de ańos), luego se observa una radiación
(generación de nuevas especies) muy rápida hacia el
Cretácico medio (hace 100 millones de ańos) y, hacia
finales de este período y comienzos del Paleógeno
(65-60 millones de ańos), el grupo ya dominaba la
mayoría de los ambientes y presentaba una
distribución cosmopolita (en toda la Tierra). Las
evidencias fósiles sugieren que este grupo de
plantas se originó en latitudes bajas, próximas al
Ecuador, desde donde migraron en dirección a ambos
polos. |
Pero... ż De dónde se originaron las flores ?
Esta es una
de las preguntas más famosas en las ciencias de la vida, que
aún es tema de debate. Charles Darwin en 1859 definió esta
pregunta como un "abominable misterio", ya que no se
encontraban indicios que permitieran explicar cómo y a
partir de que plantas se habrían originado las flores. Hoy,
con la suma de nuevas evidencias científicas sabemos muchas
cosas para explicar de donde se generaron las flores.
El registro
fósil demuestra que hubo muchos intentos en la historia
evolutiva por generar plantas que se asemejaron de una u
otra forma a las angiospermas. Algunos linajes primitivos
crearon estructuras similares a flores (como las
Pteridospermas y las Bennettitales, grupos de plantas hoy
extintos). A pesar de que estos grupos tuvieron éxito en su
época (fueron dominantes o se dispersaron por todo el
mundo), en algún momento se extinguieron y, en la
actualidad, no tenemos ningún representante para conocerlo
en su forma viva.
Fue a
partir de un ancestro vinculado a estos grupos de plantas
primitivas, que en el Cretácico Inferior o incluso antes, un
grupo minoritario de pequeńas plantas herbáceas, asociadas a
ambientes acuáticos, tuvieron la chance de diversificarse y
expandirse, y evolucionaron convirtiéndose en el gran grupo
que conocemos como angiospermas.
El registro
fósil da cuenta que las primeras angiospermas tenían flores
muy pequeńas y no tan vistosas. También nos muestra que ya
desde sus orígenes (hace 140 millones de ańos) sus flores
presentaban adaptaciones para la dispersión del pólen tanto
por el viento como por medio de sus aliados, los insectos.
Es muy probable que su íntima relación con los insectos les
haya brindado a las angiospermas una ventaja adaptativa por
sobre otros grupos de plantas.
żHay fósiles de flores en Argentina?
Si, hay
muchos fósiles de flores, como así también de frutos,
semillas, hojas, leńos, polen, etc. que dan cuenta de la
abundancia de las angiospermas en las comunidades vegetales
del pasado. En Patagonia, tenemos un excepcionalmente rico y
diverso registro fósil de angiospermas que nos permite ir
reconstruyendo la historia evolutiva del grupo en el sur del
Continente. Los restos fósiles más abundantes consisten en
polen y hojas. El registro más antiguo de angiospermas en
Patagonia, y uno de los más antiguos del Hemisferio Sur, ha
sido encontrado en la provincia de Santa Cruz y tiene
aproximadamente 120 millones de ańos, reconociéndose gracias
al intrincado patrón de venas que tienen sus hojas. Para
este momento, las plantas con flor eran un componente menor
en la comunidad vegetal, donde dominaban los helechos y
otros grupos de plantas con semilla. Cerca de 20 o 25
millones de ańos después, las angiospermas se vuelven un
componente importante de las paleo-comunidades vegetales;
aspecto que se acentuó hacia el Cretácico tardío.
|
 |
Otro registro fósil de angiospermas abundante en
Argentina lo constituye la madera petrificada. No
solo existente en aquellos bosques petrificados tan
famosos en Patagonia, sino también diseminado entre
los cantos rodados de la estepa. Al ser tan duros
resisten mucho la abrasión por los ríos, los vientos
y las condiciones climáticas extremas del invierno y
el verano quedando mezclado entre las rocas más
duras. Las flores, por ser tan delicadas, son menos
comunes en los yacimientos fósiles. No obstante
existen en Patagonia yacimientos excepcionales,
donde la fina ceniza volcánica de erupciones pasadas
sepultó estos órganos y permitió su preservación.
|
Los
yacimientos fósiles en Río Negro, Chubut y Santa Cruz
demuestran que hace 50 millones de ańos en la Patagonia se
desarrollaban selvas y bosques frondosos con plantas con
flores y frutos tan variados como en las selvas misioneras y
brasileras. Muchos de estos fósiles pueden ser vistos en la
sala de exhibición del Museo Paleontológico Bariloche.
Considerando que los cereales (arroz, maíz, trigo), pasturas
y las plantas que tienen frutos tienen flores y, por ende,
son angiospermas; podemos darnos cuenta que este grupo de
plantas constituye hoy la parte más importante de los
recursos alimenticios sobre la Tierra. Esto último, en forma
directa e indirecta, mediante la alimentación de aquellos
animales herbívoros que son fuente de los carnívoros.
Es muy
difícil pensar en un mundo con los mismos animales pero sin
la existencia de plantas con flores. No obstante esto
ocurrió por más de 350 millones de ańos en la historia
evolutiva de las plantas sobre la Tierra.
Entender
como eran las cadenas alimenticias de entonces, manteniendo
o inclusive superando la biomasa vegetal y animal en la
Tierra es todo un reto para los científicos. Quizá, de
alguna manera, su estudio pueda brindar claves para el
desarrollo de formas alternativas o de sobre-vivencia ante
climas totalmente diferentes a los actuales.
Bicentenario del Museo
Argentino de Ciencias Naturales “Bernardino Rivadavia” de
Buenos Aires, Argentina.
El
origen del Museo Argentino de Ciencias Naturales se remonta
al ańo 1812, oportunidad en la que el Primer Triunvirato,
por inspiración de Bernardino Rivadavia, invitó a las
provincias a reunir materiales para "dar principio al
establecimiento en la Capital de un Museo de Historia
Natural". Esta iniciativa se concretó en 1823, mediante una
resolución firmada por Rivadavia, entonces ministro de
Martín Rodríguez.
A lo
largo de su larga vida el Museo se alojó sucesivamente en
las celdas altas del Convento de Santo Domingo, en la
Manzana de las Luces y algunos edificios de la plazoleta
Monserrat, hasta instalarse definitivamente en 1937, en el
edificio que hoy ocupa, construido de acuerdo a los cánones
arquitectónicos vigentes en la época para los museos de
ciencias europeos.
Por sus
salas y laboratorios pasaron investigadores de la talla de
Alcides d’Orbigny Fernando Lahille, L.Kraglievich, Eduardo
Ladislao Holmberg, María Isabel Hylton Scott, Alberto
Castellanos, Rita Schiappelli, Rosendo Pascual, Enrique
Balech y Esteban Boltovskoy; y tuvo por directores a
personalidades como Herman Burmeister, Florentino Ameghino,
Angel Gallardo, Martín Doello-Jurado, Agustín Riggi,
Maximiliano Birabén, José María Gallardo, Wolfgang
Volkheimer y actualmente Edgardo J. Romero.Las colecciones
del Museo estuvieron integradas en un principio por
elementos heterogéneos que luego fueron agrupándose en forma
temática y desprendiéndose para dar origen a otros museos de
la capital. En 1947 el Poder Ejecutivo dispuso la
transferencia de las secciones: Arqueología, Etnografía y
Antropología al Museo Etnográfico dependiente de la
Universidad de Buenos Aires.
1812.
Circular cursada por el Primer Triunvirato, el 27 de junio, por
inspiración de Rivadavia, invitando a los comandantes del
interior a participar en el acopio de materiales para ‘dar
principio al establecimiento en la Capital de un Museo de
Historia Natural".
1814. En la
Gaceta Ministerial del Gobierno de Buenos Aires del día 11 de
junio, se publica el decreto por el cual se acepta la donación
efectuada por el presbítero Bartolomé Muńoz de objetos e
instrumentos de Historia Natural para "empezar a formar un
gabinete".
 |
1823. Por resolución
firmada por Rivadavia, ministro de Gobierno de Martín
Rodríguez, fechada el 31 de diciembre, se ordena adoptar
las medidas, "...que conduzcan a acelerar el
establecimiento del Museo del País, especialmente en
todos los ramos de la historia natural."
1826. El 1° de abril se
nombra a Carlos Ferraris encargado de los objetos de
Historia Natural y se instala el museo naciente en una
de las celdas altas del Convento de Santo Domingo.
|
1828. El 4
de junio Ferraris eleva al gobierno el primer inventario de
existencias y gastos invertidos en instalación y conservación.
1830. En el
curso de ese ańo visita el Museo el viajero y estudioso francés
Arsenio Isabelle y deja una descripción del mismo afirmando que
en él "se podría dictar un curso completo de historia natural".
1842. El 3
de febrero se concede licencia a Carlos Ferraris, a su pedido,
para ausentarse a Europa y es designado para reemplazarlo -
también a su propuesta - Antonio Demarchi, quien se mantiene al
frente de la dirección hasta la caída de Rosas.
1854. El 6
de mayo se designa encargado del Museo a don Santiago Torres, y
se constituye la Sociedad de Amigos de la Historia Natural del
Plata. El Museo es instalado en el edificio de la Universidad de
Buenos Aires, situado en la calle Perú en la llamada Manzana de
las Luces.
1856. El
Secretario de la Asociación Amigos de la Historia Natural del
Plata, don Manuel Ricardo Trelles, eleva un estado del Museo con
inventario de su acervo.
1857. El
sabio alemán Herman Burmeister, pasa fugazmente por la ciudad de
Buenos Aires, visita en los primeros días de febrero el Museo y
asienta su impresión sobre el mismo.
1862. Por
decreto firmado por Bartolomé Mitre, gobernador de Buenos Aires,
refrendado por su Ministro de Gobierno, Domingo Faustino
Sarmiento, del 21 de febrero, es designado Director General del
Museo Público de Buenos Aires, don Herman Burmeister.
Burmeister inicia la publicación de los "Anales" del Museo.
 |
1870. Por sugerencia de Burmeister se crea la Academia
de Ciencias de Córdoba.
1876. Se inicia la publicación de la "Descriptión
Physique de la Republique Argentine" cuya parte final
concluirá en 1886.
1884. Obtenida la capitalización de la Ciudad de Buenos
Aires, por decreto del 1° de octubre se dispone la
nacionalización del Museo Público.
1884. A raíz de aquella nacionalización se crea el Museo
de La Plata sobre la base de las colecciones de
Francisco P. Moreno.
1889. El 24 de mayo,
la Municipalidad de Buenos Aires crea el Museo Histórico
de la Capital, y el Museo Público le transfiere el
material respectivo que posee |
1892. El 2
de mayo, por causa de un accidente ocurrido en el curso de sus
trabajos en el Museo, fallece Herman Burmeister, después de
regir durante treinta ańos la institución.
1892. El 18
de abril el Poder Ejecutivo nombra director reemplazante al Dr.
Carlos Berg.
1894. Por
insistencia de su director es remodelado substancialmente el
local de la calle Perú.
1897. Entre
el 12 de mayo y el 10 de diciembre, por ausencia temporaria del
Dr. Berg, es designado director suplente el seńor Angel
Gallardo.
1902. El 19
de enero fallece el Dr. Carlos Berg.
1902. Por
decreto del 19 de abril es designado titular de la dirección el
paleontólogo Florentino Ameghino, primer director argentino del
Museo.
1910.
Durante los festejos del Centenario el Museo permanece cerrado
"por reformas".
1911. El 6
de agosto fallece el doctor Florentino Ameghino después de una
fructífera gestión científica.
1911. El 12
de septiembre es designado para reemplazarlo, el doctor Angel
Gallardo, quien se encuentra en París.
 |
1912. A su regreso de
Parías, el doctor Angel Gallardo asume la dirección del
Museo el 15 de abril.
1914. Ante los reclamos
del doctor Gallardo sobre la estrechez del local de la
calle Perú, el gobierno le cede unos edificios fiscales
en la plazoleta Monserrat que le permiten redistribuir
oficinas y colecciones.
1916. Por haber sido
designado Gallardo presidente del Consejo Nacional de
Educación, se nombra director interino al Secretario
Agustín G. Péndola.
1919. Por resolución
ministerial de fecha 11 de agosto, se transfiere la
dirección interina al jefe de sección Carlos Ameghino.
|
1923. Por
decreto fechado el 28 de diciembre, firmado por el presidente
Marcelo T. de Alvear, refrendado por el ministro Sagarna, se
denomina al Museo con el nombre de su fundador: Bernardino
Rivadavia, y se dispone festejar el centenario de su creación.
1923. Por
el mismo decreto del 28 de diciembre, se nombra director titular
al profesor Martín Doello-Jurado, quien asume el cargo en el
acto que se celebra como rememoración del centenario del Museo
el Día 31 de diciembre.
1925. El 31
de diciembre, se coloca la piedra fundamental del edificio a
construir por la Dirección General de Arquitectura de la Nación,
en el Parque Centenario.
1928. Por
iniciativa de Doello-Jurado se inaugura la Estación
Hidrobiológica de Puerto Quequén, dependiente del Museo.
1929. Se
inaugura la primera sección del nuevo edificio.
1937.
Concluido el edificio en su estado actual, el Museo abandona
definitivamente los viejos locales de la Manzana de las Luces y
de la Plaza Monserrat e instala con mayor amplitud y propiedad
sus colecciones.
1946. Por
renuncia del profesor Martín Doello-Jurado, por decreto fechado
el 10 de julio se nombra su reemplazante al doctor en Ciencias
Naturales Agustín E. Riggi.
1947. A
pedido de la dirección, el Poder Ejecutivo dispone por decreto
dictado el 10 de junio, la transferencia de las secciones:
Arqueología, Etnográfica y Antropológica a la Universidad de
Buenos Aires, con destino al Museo Etnográfico.
 |
1948. Por decreto del 4 de diciembre, se crea, anexo al
Museo, el Instituto Nacional de Investigación de las
Ciencias Naturales.
1955. Sobrevenido el movimiento revolucionario de
setiembre, se designa interventor del Museo al doctor
Adolfo Dago Holmberg. |
1958.
Elegidas las autoridades constitucionales, renuncia el doctor
Holmberg y asume la dirección interina el Director General de
Cultura, ingeniero José Babini, designado por resolución
ministerial del 26 de mayo, cesando el 12 de agosto de 1958.
1958. En la
misma fecha es designado director, también con carácter
interino, el doctor en Ciencias Naturales Mario Egidio Teruggi y
cesa en su mandato el 7 de setiembre de 1959.
1959. Por
decreto del Poder Ejecutivo del 26 de agosto se nombra director
titular al doctor en Ciencias Naturales Max Birabén quien asume
el cargo el 7 de septiembre.
1972.
Después de doce ańos de acción efectiva y provechosa, por
razones de salud, el 31 de mayo, renuncia el doctor Birabén.
1973. El
licenciado en Ciencias Naturales José María A.F. Gallardo, quien
había sido designado a cargo de la Dirección el 6 de octubre de
1971 por enfermedad del doctor Birabén, es designado titular,
por concurso, el 17 de abril.
1974. Por
resolución del 6 de diciembre, la Inspección General de Personas
Jurídicas, concede la personería solicitada por la "Asociación
Amigos del Museo Argentino de Ciencias Naturales Bernardino
Rivadavia, constituida el 29 de agosto de 1969.
1986. El
Museo deja de depender de la Dirección Nacional de Museos para
hacerlo de la Secretaría de Ciencia y Tecnología (SECyT), que en
ese momento dependía directamente de la Presidencia de la
Nación.
 |
1994. Fallece el Dr. José María A.F. Gallardo el 12 de
octubre, estando a cargo de la Dirección del Museo como
Investigador Principal del CONICET. Antes de finalizar
el ańo es designado a cargo de la Dirección el Dr.
Wolfgang Volkheimer, Investigador Superior del CONICET,
especialista en palinología y paleobotánica.
1996. El Decreto del
Poder Ejecutivo de la Nación 1274/96 del 7 de noviembre
dispone que el Museo sea transferido desde la Secretaría
de Ciencia y Tecnología al CONICET (Concejo Nacional de
Investigaciones Científicas y Tecnológicas), organismo
descentralizado dependiente de dicha Secretaría del
Ministerio de Cultura y Educación. |
1999. El
CONICET llama a concurso para ocupar el cargo de Director del
Museo y el 2 de noviembre el Presidente del CONICET pone en
funciones al Dr. Edgardo Juan Romero, Investigador Principal y
Profesor de la Universidad de Buenos Aires, que estaba
provisoriamente a cargo de la Dirección del Museo desde hacía un
ańo. En julio el MACN comienza ha estar presente en Internet por
medio de esta página y gracias al apoyo de la Red Científica y
Tecnológica (RedCyT) de la SECyT.
2002. Se
instala y pone en funcionamiento una red estructurada de
telefonía y datos que barca todas las instalaciones del Museo
permitiendo el acceso a todo el personal a los servicios de
correo electrónico (brindado por el propio servidor del Museo) e
Internet, gracias a la colaboración del Centro de Cómputos del
CRIBABB (Centro Regional del CONICET en Bahía Blanca), RETINA
(Red Teleinformática Académica) y la Fundación Antorchas.
El actual
edificio fue construido de acuerdo a los cánones arquitectónicos
y conceptos museológicos vigentes en la primera mitad de este
siglo. Esto lo convierte, aún hoy, en uno de los pocos Museos
argentinos que cuenta con un edificio concebido para su función
específica.
Los buhos,
símbolos de la sabiduría, que flanquean las ventanas del primer
piso y las arańas de bronce en sus telas que adornan las
puertas, son sólo algunos de los numerosos motivos decorativos
de interés, que el visitante puede descubrir durante su visita.
 |
El
presente edificio es solo una tercera parte del proyecto
original, y muestra en los detalles decorativos y
ornamentales temas basados en la flora y fauna
autóctonas.
Todos estos artistas fueron renombrados escultores
argentinos, algunos de ellos como Alfredo Bigatti y
Donato Proietto con abundante actuación en el país y en
el extranjero, donde merecieron destacados premios.
Se
puede observar la balaustrada de la escalera que
reproduce en hierro forjado caracoles de tierra
estilizados, y una escultura que representa unos monos
trepando a un tronco Podemos apreciar, ahora desde
adentro, las arańas en sus telas que decoran las
puertas, motivo que también se repite en las rejillas de
aereación. |
Sobre las
puertas interiores que dan acceso a la Sala de Mineralogía y
Geología y al Acuario, hay altorrelieves que muestran
respectivamente a un perezoso y a un puma. Las ménsulas que
sostienen las vigas tienen forma de murciélagos.
Preside el
vestíbulo un busto de Bernardino Rivadavia, fundador del Museo.
Fuente
Principal:
www.macn.secyt.gov.ar/
 Ver mas Artículos de Divulgación Paleontológica Ver mas Artículos de Divulgación Paleontológica |