|
Fósiles del Plioceno
de Argentina:
El
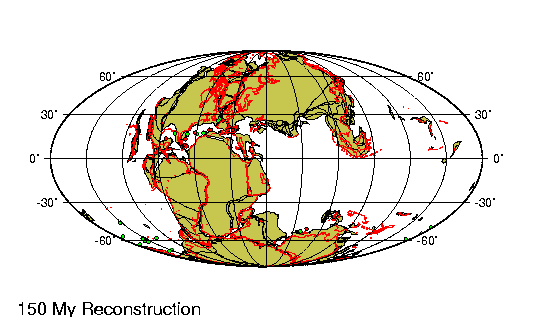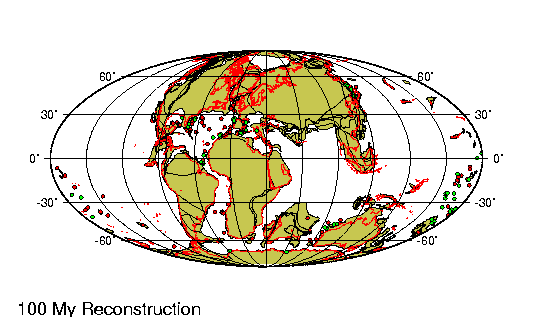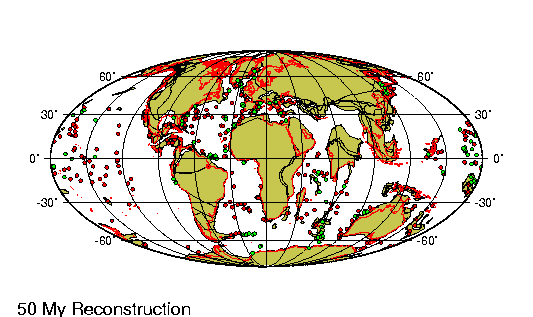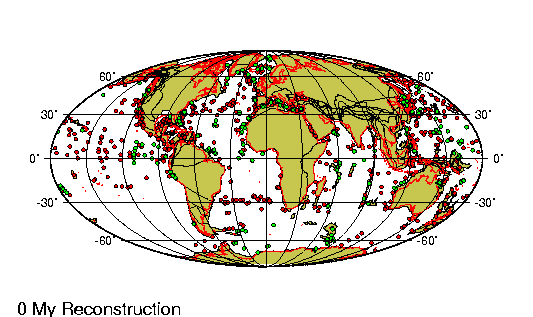
Periodo/Epoca Plioceno comprende entre los últimos 5 y 2,5 millones
de ańos de la tierra. En la Republica Argentina se encuentra
dividido en tres edades principales.

Sedimentos del Plioceno de Miramar.
|
 |
La
edad mamífero "Montehermorense" comprende un lapso
intermedio entre el Mioceno tardío y el Plioceno temprano.
Tiene una antigüedad entre 6 y 4 millones de ańos. Los
depósitos sedimentarios afloran a unos 15 kilómetros al sur
de la localidad de Pehuen-co, Provincia de Buenos Aires,
Argentina. La edad "Chapadmalalense" es tal vez la más
importante del Plioceno por la abundancia de restos fósiles
y evidencias ambientales. Tiene entre 4 y 3,2 millones de
ańos y corresponden a sedimentos del litoral marítimo
pampeano, ubicados entre la ciudad de Mar del Plata y
Miramar, Provincia de Buenos Aires. Durante el Final del
Plioceno ocurrieron tres fenómenos naturales, los cuales
decidieron la suerte del 80 % de las especies
endémicas y autóctonas. Una de ellas fue el Gran Intercambio
Biótico Americano, que fue la migración de los mamíferos al
restablecerse la unión entre América del Norte y Sur,
provocando la llegada de la fauna invasora. Otro de los
fenómenos fue el impacto de un Asteroide, el cual ocurrió
hace 3,3 millones de ańos. Ha esto se le suma, hacia el
final del Plioceno, una estrella del grupo de estrellas O y
B de la Asociación estelar de Scorpius-Centaurus a unos 380
a 470 ańos luz de la Tierra, explotó como supernova, lo
suficientemente cerca de la Tierra como para provocar un
gran deterioro en la capa de ozono, lo que pudo haber sido
la causa de una extinción masiva en los océanos. Para ello
se basaron en las anomalías del isótopo de esa época
encontradas en los fondos oceánicos. <<
Principales sitios
de Argentina. |
|
Recientemente se incluyó la edad
"Marplatense", la cual se encuentra dividida en tres sub-edades
(sanandresense, vorohuense y barrancalobense) que tienen una
antigüedad entre 3,2 y 1,9 millones de ańos, ubicadas
principalmente al sur de la ciudad de Mar del Plata hasta
Miramar. En el continente se produjo un evidente
desplazamiento de distintos grupos de vertebrados hacia
condiciones más favorables existentes al norte de Patagonia.
A partir del Plioceno el registro fósil pertenece
exclusivamente a algunas localidades del norte y centro del
territorio argentino, a diferencia de las anteriores (del
Mioceno al Paleoceno) que se encontraban principalmente en
Patagonia. Existía un gradiente decreciente de humedad en
dirección al oeste. En el territorio se desarrollaron las
llanuras, con extensas praderas bajo un clima templado -
cálido y húmedo. Ingresaron provenientes de Norteamérica
numerosas especies de carnívoros placentarios.
Posteriormente al elevarse las Sierras Pampeanas se produce
una "sombra de lluvias" al este de las mismas, con lo cual
se produjo la desertización de las áreas sub-andinas. Este
periodo coincide con un deterioro en el clima, con un
enfriamiento a nivel global, fenómenos de glaciaciones, un
aumento en el gradiente térmico latitudinal, y un descenso
en el nivel del mar, lo que provoco la disminución de
especies. |
Mesembriornis
milneedwardsi.
Moreno, 1889.
|
Tizziana
Magnussen, con el cráneo y pico
Mesembriornis milneedwardsi
en el Museo de Ciencias Naturales de Miramar. |
Ave
Carnívora.
Como ya se había aclarado
anteriormente, las aves carnívoras, tanto corredoras como voladoras,
alcanzaron durante el Terciario dimensiones extraordinarias y se
convirtieron en los principales depredadores debido a la ausencia de
mamíferos competidores de gran tamańo. Mesembriornis
milneedwardsi fue un ave corredora y carnívora que superaba los 1,8
metros de altura. Sus restos fósiles fueron hallados por primera vez
en la Farola de Monte Hermoso (Montehermosense–Chapadmalalense) por
Moreno y Mercerat en 1891, y depositados en el Museo de La Plata.
Posteriormente, en 1908, se colectó un esqueleto casi completo
procedente de la misma zona, actualmente preservado en el Museo
Argentino de Ciencias Naturales de Buenos Aires. La punta del pico
dirigida hacia abajo, similar a la de las aves rapaces, revela
hábitos carnívoros; sus principales presas habrían sido animales de
talla media y pequeńa, como Paedotherium o
Pseudotypotherium, a los que perseguía hasta alcanzarlos
para golpearlos con el pico y, una vez detenidos, sujetarlos con sus
poderosas patas, clavando las garras hasta someterlos por completo.
No desaprovechaba la carrońa, especialmente durante períodos de
escasez. Sus alas eran muy reducidas y con musculatura atrofiada, lo
que le impedía volar. El cráneo superaba los 45 centímetros de
longitud. Se extinguió durante el Plioceno, hace unos 3 millones de
ańos, sin dejar representantes actuales; su nicho ecológico fue
luego ocupado por numerosos mamíferos placentarios que ingresaron al
continente tras la formación del istmo. Algunos restos proceden de
sedimentos de Monte Hermoso y Barranca de los Lobos. La especie
Hermosiornis rapax podría corresponder a Mesembriornis milneedwardsi
según algunos estudios. Géneros y especies
relacionados: Psilopterus
colzecus, Palaeopsilopterus itaboraiensis, Chunga incerta,
Hermosiornis australis y
Hermosiornis rapax.
Psiloterus s.p.
Moreno y Mercerat, 1891.
| |
|
|

Cráneo de Psiloterus sp
en el Museo de Cs Nat. de Miramar.
|
|
 |
Ave Carnívora.
Es un género
extinto de forusrácidos (“aves del terror”) que vivió desde mediados
del Oligoceno hasta fines del Mioceno en Argentina. En comparación
con otros forusrácidos, sus representantes eran relativamente
pequeńos y gráciles. Un cráneo hallado en las cercanías de la ciudad
de Miramar pertenece a uno de los últimos integrantes de la
subfamilia Psilopterinae y procede del Plioceno tardío, hace unos 3
millones de ańos, correspondiente a la Edad Chapadmalalense de la
provincia de Buenos Aires, aunque los registros más antiguos del
género provienen del Mioceno temprano–medio (Santacrucense) de
Patagonia. El cráneo conserva una longitud aproximada de 23
centímetros sin el pico. A pesar de no encontrarse completo y haber
sido sepultado en condiciones ambientales desfavorables, presenta
una característica excepcional: la preservación de la esclerótica
ocular, formada por delicados huesecillos ubicados en la órbita.
Asimismo, se observan marcas atribuibles a un par de caninos en su
superficie superior. Esta ave habría alcanzado una altura cercana a
1,6 metros. En la actualidad existen representantes emparentados con
esta familia en Argentina: los dos únicos miembros de la familia
Cariamidae, la chuńa de patas rojas (Cariama cristata),
de unos 95 cm de altura, y la chuńa de patas negras (Chunga
burmeisteri), de aproximadamente 78 cm. Su dieta incluía
principalmente pequeńos mamíferos como Paedotherium o
Dasypus, aunque existía una amplia diversidad de
presas disponibles en ese momento. Especies y especies
relacionadas: P. bachmani, P.
lemoinei, P. affinis y P. colzecus.
Heterorhea dabbenei.
Rovereto, 1914.
 |
 |
|
Pata recreada completa de un Rheiforme,
en el Museo de Ciencias Naturales de Miramar. |
Aspecto
de Heterorhea. Por MarMag.2025. |
Ave.
Rheiformes.
Se trata de un ave del orden Rheiformes, nombre que
reciben dos aves sudamericanas similares al avestruz, emparentadas con
el género Rhea del Pleistoceno y con el ńandú actual. Era
de mayor tamańo que los representantes actuales de Sudamérica y, al
igual que ellos, poseía tres dedos en cada pata. La cabeza y el cuello
estaban completamente cubiertos de plumas. La cola no se encontraba
desarrollada, pero presentaba largas plumas que colgaban y cubrían la
parte posterior del cuerpo. Probablemente incubaban los huevos de manera
similar a las formas actuales, en las que varias hembras utilizan un
mismo nido y un solo macho puede incubar hasta 50 huevos, aunque este
comportamiento no puede confirmarse con el registro fósil.
Heterorhea era un corredor veloz gracias a sus largas patas y a
la proporción de sus huesos, que le permitían alcanzar altas
velocidades. Es posible que se desplazara en grupos familiares, salvo
durante la época reproductiva, cuando se congregaría en grandes
bandadas, volviéndose más vulnerable frente a depredadores marsupiales.
Su dieta habría sido variada, incluyendo pastos, insectos y frutos.
Aunque se trata de un género poco frecuente, algunos restos proceden del
Terciario bonaerense y tucumano, representados por huesos aislados y
cáscaras de huevo atribuidas tentativamente a Heterorhea.
El primer registro, en 1914, se basó en un tarsometatarso proveniente de
la Formación Monte Hermoso, y posteriormente se describió un nuevo
género y especie a partir de un fémur hallado en los mismos niveles de
los barrancos de Monte Hermoso (provincia de Buenos Aires). Estos
constituyen los únicos registros pliocenos conocidos y corresponden a
formas más grandes y robustas que los géneros actuales de Rheiformes.
Géneros y especies relacionados: Heterorhea
dabbenei, Rhea anchorenensis, Rhea americana, Rhea fossilis y
Pterocnemia pennata.
Nothura parvula.
Tambussi, 1989.
|
Restos fósiles del humero y
sacro de un Nothura s.p, del Plioceno de Miramar.
Museo de Ciencias Naturales de Miramar y reconstrucción por
MarMag.2025. |
Ave,
Ratites.
Los tinámidos son aves terrestres cuyo
registro fósil se conoce principalmente en el Plioceno y Pleistoceno
de Argentina. La familia Tinamidae es endémica de la región
Neotropical y, vulgarmente conocidos como inambúes, se caracterizan
por su escasa capacidad de vuelo y hábitos terrícolas. Si bien se
han hallado restos fósiles en Brasil y Perú, la mayor parte del
registro procede de Argentina, donde el resto más antiguo fue
descubierto en el Mioceno tardío de la provincia de La Pampa. Los
tinamúes pertenecen a los Paleognathae, tradicionalmente divididos
en dos linajes: tinamúes y ratites; a diferencia de estas últimas,
los tinamúes conservaron la capacidad de volar. Probablemente
presenten un aspecto similar al de los ancestros voladores de las
ratites. Estas últimas se distribuyen en Nueva Zelanda (kiwis),
África (avestruces), América (ńandúes) y Australia y Nueva Guinea (emúes
y casuarios), mientras que los tinamúes son exclusivos del
Neotrópico. Las relaciones filogenéticas dentro de los paleognatos
han sido debatidas durante décadas, existiendo consenso únicamente
sobre la monofilia del grupo, la de los tinamúes y la del clado emú-casuario.
Respecto a su radiación evolutiva, se proponen dos hipótesis: una
sugiere una diversificación gondwánica durante el Cretácico, con la
posterior deriva continental explicando la distribución actual; la
otra plantea una radiación posterior al evento de extinción
Cretácico–Paleógeno, a partir de ancestros voladores, de los cuales
sólo los tinamúes conservaron esa capacidad. El género Nothura
Wagler, 1827 incluye actualmente cinco especies que habitan
ambientes abiertos con parches de vegetación arbórea en Sudamérica.
Solo se conocen dos especies fósiles: Nothura parvula
Tambussi, 1989 del Plioceno, y Nothura paludosa
Mercerat, 1897 del Pleistoceno, ambas halladas en la provincia de
Buenos Aires.
Vultur messii.
Degrange, F. et al. 2023.
|
 |
|
 |
|
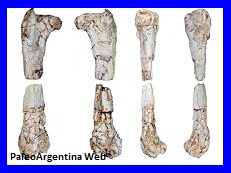
Femur de Vultur messii,
del Plioceno de Catamarca. |
|
Aspecto del cóndor
Vultur messii. Por Santiago Druetta. |
Ave. El registro fósil de cóndores (Aves,
Cathartiformes) en América del Sur es escaso y, particularmente en
Argentina, la mayoría de los restos asignados a este grupo han sido
objeto de debate. La presencia de una especie fósil del género
Vultur, recuperada de depósitos de casi 5 millones de ańos de
antigüedad, coincide con el levantamiento de los cerros pampeanos y
de los Andes, así como con el inicio de la aridificación regional.
La existencia de cadenas montańosas incipientes sugiere que el
centro de origen de este género estaría vinculado a la cordillera de
los Andes, lo que implica una llegada temprana y una posterior
diversificación de los cóndores en Sudamérica. En la actualidad, el
género Vultur está representado únicamente por
Vultur gryphus, el cóndor andino, considerado el ave
voladora más grande del mundo por la combinación de un peso cercano
a los 15 kg y una envergadura de hasta 3,3 metros. Es principalmente
carrońero y prefiere cadáveres de gran tamańo, como los de ciervos o
ganado. Alcanza la madurez sexual entre los cinco y seis ańos y
nidifica en formaciones rocosas inaccesibles, a altitudes de entre
3000 y 5000 metros sobre el nivel del mar. La posición taxonómica
exacta del cóndor andino y de las restantes especies de buitres del
Nuevo Mundo continúa siendo debatida, ya que, aunque similares en
apariencia y función ecológica a los buitres del Viejo Mundo,
evolucionaron de ancestros distintos. Una nueva especie fósil fue
hallada en afloramientos geológicos del noroeste argentino y
homenajea al futbolista Lionel Messi, a casi un ańo de la histórica
obtención de la Copa del Mundo.
Ceratophrys ameghinorum.
Fernicola, 2001.
|
Esqueleto de
un Ceratophrys ornata viviente en el MACN.
Cráneo de
Ceratophrys ameghinorum y reconstrucción en vivo.
Museo de Ciencias Naturales Punta Hermengo de
Miramar. |
Anfibio. Anura.
Los escuerzos del género
Ceratophrys conforman un peculiar grupo de anuros
neotropicales caracterizados por su notable robustez corporal y una
cabeza desproporcionadamente grande. Actualmente se reconocen seis
especies vivientes asignadas a este género, de las cuales solo dos
se distribuyen en la República Argentina. En las regiones chaqueńa y
central del país se encuentra Ceratophrys cranwelli
Barrio, 1980, mientras que Ceratophrys ornata Bell,
1841 restringe su distribución a la región pampeana. Estos animales
poseían una cabeza ancha, hidrodinámica y cubierta por ásperas
molduras óseas; las mandíbulas eran amplias y semicirculares,
provistas de una hilera de pequeńos dientes aserrados, con ocho
dientes premaxilares y entre 30 y 40 maxilares, mientras que la
mandíbula inferior carecía de dentición pero presentaba un borde
extremadamente afilado. Excavaban cuevas utilizando tubérculos
córneos de bordes agudos presentes en las patas posteriores.
Probablemente se desplazaban con torpeza en tierra firme, pero en el
agua eran rápidos y eficientes depredadores, capaces de capturar
pequeńas aves, mamíferos, insectos, peces e incluso individuos
juveniles de su propia especie. El registro fósil de este grupo es
muy escaso; sin embargo, recientemente se realizaron hallazgos
significativos al norte de la ciudad de Miramar, incluso dentro de
una crotovina junto a otros vertebrados del Plioceno superior. El
resto más antiguo de la subfamilia Ceratophryinae corresponde a
W. gerholdi del Mioceno medio de Ingeniero Jacobacci,
noroeste de Río Negro, considerado afín al género Ceratophrys.
Recién en el Plioceno tardío reaparecen registros fósiles de esta
subfamilia en el sur de la provincia de Buenos Aires (Monte Hermoso,
Formación Montehermosense). Los primeros hallazgos fueron realizados
por Ameghino en 1899, quien los asignó a Ceratophrys prisca.
Durante el Plioceno se registra Ceratophrys sp. tanto
en el noroeste argentino (Catamarca) como en el sur bonaerense. La
fauna de la región subandina de este período presentaba un marcado
carácter subtropical, lo que sugiere condiciones ambientales
similares a las actuales del oeste del distrito chaqueńo, que
aparentemente también se extendían hacia el sudoeste bonaerense. El
levantamiento final de las Sierras Pampeanas durante el Plioceno
tardío habría bloqueado los vientos húmedos del nordeste, provocando
la desertificación de las áreas occidentales. En el Piso
Chapadmalalense se registran ejemplares diploides en Miramar, Punta
Lobería y Punta Vorohué, y tetraploides en Quequén Salado y
Chapadmalal. En el Piso Barrancalobense, hacia el final del
Plioceno, se documenta un fósil diploide. Durante el Pleistoceno, a
lo largo de la costa atlántica, se hallaron nuevos restos asignados
a Ceratophrys (Piso Ensenadense). También se conocen materiales del
Pleistoceno superior de Bolivia, en Ńuapua y Tarija, así como restos
coetáneos de Brasil, en Lagoa Santa, Minas Gerais.
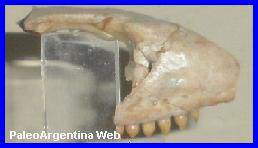
Salvator (Tupinambis) teguixin.
Daudin, 1802.
|
 |
|
 |
|
Fragmento de
cráneo y maxilar de Tupinambis exhibido en el Museo de
Ciencias Naturales
de Miramar. |
|
Reconstrucción en vivo del genero Tupinambis teguixin
del Plioceno pampeano.
Por MarMag.2025. |
Reptil
Squamata.
Es el lagarto de mayor
tamańo de la familia Teiidae, registrado desde el Mioceno temprano de
Patagonia y con presencia continua en Argentina durante todo el Mioceno,
extendiéndose hasta el Holoceno. Los reptiles escamosos (Squamata)
constituyen el segundo grupo más numeroso de vertebrados terrestres, con
más de 8200 especies, superados únicamente por las aves; sin embargo,
pese a su actual diversidad y abundancia, han sido tradicionalmente
relegados en la literatura paleontológica. En la actualidad, el género
Tupinambis, hoy reconocido como Salvator sp.,
está representado por al menos seis especies: T. duseni, T.
longilineus, T. merianae, T. quadrilineatus, T. rufescens y T. teguixin,
todas restringidas al territorio sudamericano al este de los Andes.
Recientemente fue descripta una forma extinta, T. uruguaianensis,
procedente del Pleistoceno de Brasil, de mayor tamańo que cualquiera de
las especies actuales. En 1914, Rovereto describió cuatro especies
fósiles de Tupinambis (T. preteguixin, T. prerufescens, T.
brevirostris y T. multidentatus) provenientes de Monte Hermoso,
a unos 60 km al este de Bahía Blanca, sobre la costa atlántica del sur
bonaerense. Su registro paleontológico en sedimentos pampeanos se
extiende desde el Mioceno tardío hasta el Holoceno temprano, aunque su
mayor abundancia corresponde al final del Plioceno. Se caracterizaba por
una marcada diferenciación dentaria, con incisivos, dientes caniniformes
y otros aplanados similares a molares. Podía superar el metro de
longitud total, aunque la cola representaba más de la mitad del cuerpo.
Los machos presentaban la cabeza más ancha debido al desarrollo de los
músculos mandibulares. Su dieta incluía anfibios, gusanos, reptiles y
pequeńos mamíferos, como cricétidos y algunos marsupiales. Sus restos
aparecen ocasionalmente articulados dentro de paleocuevas excavadas por
el propio animal o reutilizadas tras haber sido realizadas por otros
vertebrados. Importantes hallazgos para la paleofauna pampeana proceden
del “Uquiense” del arroyo Las Brusquitas y Barranca de los Lobos, así
como de sedimentos modernos y sitios arqueológicos. En la región
pampeana se lo registra en las formaciones Monte Hermoso, Chapadmalal y
Vorohué.
Amphisbaena sp.
Linnaeus, 1758.
|
 |
 |
 |
| Cráneo de un
Amphisbaena (*). |
Vértebra del Plioceno
encontrada cerca de Miramar. (autores). |
Aspecto de un
Amphisbaena (*). |
Reptil
Squamata.
Es un género de
reptiles anfisbénidos de la familia Amphisbaenidae, comúnmente conocidos
como culebras de dos cabezas, morronas o lagartijas gusano. Estas
especies presentan un cuerpo cilíndrico, sin extremidades, con una cola
corta y redondeada poco diferenciada del resto del cuerpo; la cabeza
también es redondeada y apenas distinguible, con ojos pequeńos
semiocultos bajo la piel y sin aberturas auditivas externas. La piel
está conformada por anillos subdivididos en pequeńas escamas
cuadrangulares. Las especies del género Amphisbaena pasan
la mayor parte de su vida bajo tierra, excavando galerías en busca de
alimento, compuesto principalmente por insectos y otros invertebrados
del suelo. Restos fósiles de Amphisbaena sp. fueron
identificados en sedimentos chapadmalalenses (Plioceno temprano–medio)
en las cercanías de la ciudad de Miramar, constituyendo hasta el momento
el registro más antiguo del grupo en América del Sur. En la región
costera de Miramar se encuentra actualmente A. angustifrons,
mientras que A. heterozonata habita áreas próximas, por lo que la
presencia del género en el Plioceno resulta coherente con su
distribución actual.
Phrynops sp.
Schweigger, 1812.
|

|
 |

|
|
Placas del caparazón de Phrynops,
del Plioceno de la prov. de Entre Ríos. Museo de La Plata. |
Cráneo de un ejemplar
actual de Tortuga de laguna del genero Phrynops. (*) |
Aspecto del Quelonio Phrynops geoffroanus.
Por MarMag.2026 |
Reptil Testudines.
El género Phrynops está
registrado desde el Mio-Plioceno de Argentina y el Mioceno superior
de Uruguay. Los antecedentes fósiles atribuidos a este género se
restringen, por el momento, a sedimentos del noroeste argentino
(Catamarca y Tucumán), a la región mesopotámica argentina y al
departamento de San José, en Uruguay. También se mencionan restos
asignables a Phrynops en sedimentos pleistocenos del
Carcarańá, provincia de Santa Fe. Desde mediados del siglo XIX, la
existencia de placas fósiles y material fragmentario fue seńalada
por Bravard, Burmeister y Ambrosetti, quienes realizaron
determinaciones específicas pese a la fragmentación del material.
Wieland, en 1923, citó para la región del Paraná a
Parahydraspis paranaensis como un nuevo taxón de pleurodiro,
aunque posteriormente Mlynarski lo consideró sinónimo de
Phrynops geoffroanus. En el Mesopotamiense se han registrado
numerosas tortugas, principalmente de la familia Chelidae y del
género Phrynops. Los quélidos, considerados excelentes
indicadores paleoambientales, sugieren condiciones subtropicales y
húmedas, con ambientes bajos, inundables y con abundante vegetación,
lo que explica la elevada diversidad de tortugas acuáticas y otros
reptiles en estas regiones.
Colubridae.
Oppel, 1811.
|
 |
 |
 |
|
Vértebras y costillas de un
Colubridae recreados in situ. Museo de Miramar. |
Vértebra del
Pleistoceno. Museo de San Pedro. Prensa. |
Aspecto del
pequeńo Colubridae. (*). |
Reptil, Serpentes.
Los
colúbridos o culebras (familia Colubridae) constituyen un amplio
grupo de serpientes caracterizadas por presentar la cabeza
recubierta por grandes escamas dispuestas de forma típica. Las
escamas dorsales y laterales del cuerpo son aproximadamente
romboidales, mientras que en la región ventral poseen una única fila
de escamas ensanchadas. Generalmente son diurnas, con ojos bien
desarrollados y pupila usualmente circular. Los registros fósiles de
serpientes en depósitos pliocenos son extremadamente escasos y
suelen consistir en elementos aislados, como vértebras o costillas,
razón por la cual resulta más apropiado referirse a la familia
Colubridae en lugar de asignaciones genéricas o específicas. El
origen de la fauna colubrídica sudamericana se ha interpretado
tradicionalmente como resultado de un ingreso pasivo desde América
del Norte, previo al establecimiento definitivo del puente
centroamericano, probablemente durante el Mioceno temprano o incluso
antes. Algunos hallazgos relevantes proceden de la Formación
Chapadmalal, en la localidad fosilífera de Las Brusquitas, a unos 5
km al norte de la ciudad de Miramar. Estos ofidios resultan de gran
importancia para la paleontología, ya que permiten realizar
inferencias paleoambientales. Al igual que sus parientes actuales,
se habrían alimentado de pequeńos vertebrados y principalmente de
insectos.
Bothrops
alternatus. Duméril, Bibron & Duméril,
1854.
|
 |
|
 |
|
Vértebras fósiles de Bothrops encontradas en el
Plioceno de Monte Hermoso, comparadas con las vértebras de
una Yarará viviente. |
|
Aspecto de Bothrops,
del Plioceno bonaerense, indistinguible de
Bothrops alternatus. Por MarMag.2025. |
Reptil, Serpentes.
Dentro de Colubroidea, el grupo
monofilético de serpientes más avanzadas, en Sudamérica están
representadas actualmente las familias Elapidae, Colubridae y
Viperidae. En relación con esta última, en ańos recientes se han
intensificado los estudios sobre la sistemática y evolución
biogeográfica de géneros como Crotalus, Lachesis, Bothriopsis,
Bothriechis y Bothrops. Este último incluye
serpientes de mediano a gran tamańo, de cuerpo robusto, que pueden
alcanzar alrededor de 1,5 metros de longitud y habitaron el Plioceno
bonaerense en ambientes abiertos, cerrados y de pastizal. Al igual
que sus representantes actuales, eran altamente venenosas y se
alimentaban de pequeńos mamíferos. Un equipo conjunto del LACEV, la
Fundación Azara y el Museo Municipal “Carlos Ameghino” de Mercedes
dio a conocer restos fósiles correspondientes a una serie de once
vértebras del tronco de una especie indistinguible de Bothrops
alternatus (yarará grande o víbora de la cruz). Este
hallazgo, procedente de la Farola de Monte Hermoso, en la provincia
de Buenos Aires, constituye el registro más antiguo conocido de
yararás en el continente. Hace unos 3 millones de ańos, la región
estaba habitada por manadas de megamamíferos como gliptodontes y
perezosos gigantes, junto con fauna típica de ambientes chaqueńos,
como chuńas, escuerzos, boas de las vizcacheras y pecaríes. Hacia
los 2,5 millones de ańos, el clima se volvió más frío y seco,
marcando el inicio del Pleistoceno y provocando el desplazamiento
progresivo de esta fauna hacia latitudes más septentrionales.
Estudios de ADN mitocondrial sugieren que el primer vipérido habría
ingresado a Sudamérica durante el Mioceno, entre 23 y 10 millones de
ańos atrás, utilizando un puente terrestre entre América Central y
del Sur; este ancestro, denominado Protobothrops,
habría dado origen a todas las especies actuales de Bothrops,
en concordancia con el ingreso documentado de diversos linajes de
mamíferos durante ese mismo intervalo temporal.
Trigodon gaudryi.
Ameghino, 1887.
|
|
|
|
|

|
 |

|
|
Cráneo y Mandíbula de Trigodon,
exhibido en el Museo de La Plata. Hallado en Monte Hermoso,
Prov. Buenos Aires. |
Cráneo de un genero afín,
Paratrigodon s.p. Prov. Buenos Aires. Museo de La Plata. |
Aspecto de la cabeza de
Trigodon. Por
MarMag.2025. |
Mamífero
Notoungulado.
Es el último género
gigante de los Haplodontheriinae, al menos en la región pampeana.
Fue uno de los representantes del orden Notoungulata, un grupo que
se diversificó en América del Sur colonizando una amplia variedad de
ambientes. Existió desde hace aproximadamente 11,6 millones de ańos
hasta hace 3 millones de ańos, durante el Mioceno superior y el
Plioceno. Se trataba de animales de patas y cuello cortos, que aún
no habían desarrollado una cruz en forma de joroba como los
toxodontos posteriores. Presentaban adaptaciones semiacuáticas y se
alimentaban de vegetación abundante en las orillas de cuerpos de
agua. La presencia de una gran fosa en el cráneo de este gigantesco
animal llevó a los paleontólogos a suponer que poseía un cuerno
córneo curvado, utilizado para defenderse de los carnívoros de su
época, como las grandes aves corredoras y los marsupiales de caninos
muy desarrollados. Su cuerpo era voluminoso y macizo, cubierto por
una piel gruesa. Las extremidades, cortas y robustas, no estaban
adaptadas para la carrera. Su peso se estima entre 1,5 y 2
toneladas. Los principales restos provienen de los acantilados
marítimos de la costa bonaerense y de algunos sitios del interior de
la provincia de Buenos Aires. Se extinguió a fines del Plioceno,
siendo reemplazado ecológicamente por Toxodon, muy abundante durante
el Pleistoceno, y es considerado un fósil característico de la Edad
Montehermosense. Su distribución incluyó la ladera oriental de los
Andes en Bolivia y Perú, las llanuras de Uruguay, la Formación
Solimőes en el alto río Acre (Brasil) y la provincia de Buenos Aires
(Argentina). Géneros y especies
relacionados: Toxodon chapalmalensis, Toxodon
darwini, Toxodon ensenadensis, Toxodon platensis, Toxodon gezi,
Toxodon gracilis, Xotodon prominens, Xotodon ambrosetti, Trigodon
minus, Alitoxodon vetustum, Mixotoxodon larensis y
Nonotherium hennigi.
Paedotherium typicum.
Ameghino, 1887.
|
|
|
|
|

|

|

|
|
Esqueleto
preservado en el Museo "Punta Hermengo" de Miramar. |
Cráneo y
mandíbula en el Museo de Ciencias Naturales de Miramar. |
Reconstrucción en vivo.
Por MarMag.2025. |
Mamífero
Notoungulado.
Paedotherium
fue el representante más pequeńo de un
grupo particularmente abundante. Sus dimensiones y aspecto en vida
recuerdan a la liebre patagónica o mara (Dolichotis australis).
Presentaba un cráneo corto y robusto, con cierta semejanza al de los
roedores, incisivos muy prominentes de crecimiento continuo, cortos
y cincelados, ausencia de caninos, y premolares y molares alargados,
bilobulados, seis por cada mitad del maxilar y la mandíbula. Poseía
órbitas grandes, lo que sugiere hábitos crepusculares o nocturnos,
así como un aparato auditivo muy desarrollado, indicativo de
posibles costumbres terrestres y subterráneas. Los miembros
anteriores eran digitígrados, mientras que los posteriores, más
largos, eran plantígrados. Se alimentaba de vegetación abrasiva,
dura y rugosa, y probablemente vivía en madrigueras excavadas por el
propio animal. Fue una especie muy exitosa, con una distribución
temporal que abarca desde el Mioceno hasta el Pleistoceno inferior.
Su extinción se asocia a cambios ambientales que afectaron
drásticamente a sus poblaciones. Es uno de los notoungulados mejor
conocidos de la región pampeana debido a la abundancia de sus restos
fósiles, particularmente en las edades Montehermosense y
Chapadmalalense. Algunos investigadores sostienen que
Paedotherium typicum y Paedotherium insigne
serían descendientes del representante mioceno Paedotherium
minor. Géneros y especies
relacionados: Paedotherium typicum,
Paedotherium bonaerense, Paedotherium insigne y
Tremacyllus impressus.
Xotodon catamarcensis.
Ameghino, 1887.
|
 |
|
 |
| Cráneo de
Xotodon del Plioceno de Catamarca. Ejemplar depositado en
Museum für Naturkunde, Berlin. . |
|
Posible
aspecto del toxodontido Xotodon. (*). |
Mamífero
Notoungulado. Fue
un género perteneciente a la familia extinta del suborden Toxodonta,
dentro del también extinto orden de ungulados sudamericanos
Notoungulata. El ejemplar descubierto en el Plioceno de Catamarca
correspondía a un gran herbívoro, de tamańo comparable al del actual
rinoceronte negro, con tres dedos en cada pata. La radiación
adaptativa de los ungulados paleocenos de América del Sur fue
temprana y rápida, lo que dificulta la elaboración de modelos
filogenéticos completamente explicativos. Derivados de pequeńos
herbívoros primitivos sin competidores, evolucionaron de manera
independiente y aislada del resto de los ungulados durante gran
parte del Cenozoico. Los mamíferos originarios de Sudamérica
evolucionaron en un contexto de continente-isla. Hacia 1900,
Florentino Ameghino inició un extenso catálogo que incluye
clasificaciones, estudios comparativos y descripciones de más de
9000 animales extintos, muchos descubiertos por él mismo. Los
notoungulados comprenden más de 100 géneros agrupados en cuatro
subórdenes. Los ungulados sudamericanos representan un caso de
aislamiento geográfico tan notable como el de los marsupiales
australianos. Los meridiungulados, comúnmente denominados ungulados
sudamericanos, incluyen a los toxodóntidos, animales con dientes de
coronas excepcionalmente altas y curvadas, con raíces abiertas de
crecimiento continuo que compensaban el desgaste producido por el
consumo de hierbas duras de la pampa. Otros géneros y
especies relacionados son
Pericotoxodon, Trigodon, Toxodon, Nesodon, Adinotherium,
Haplodontherium y Dinotoxodon.
Seudotypotherium hystatum.
Cabrera, 1904.
|
 |
|
 |
|
Cráneo de Pseudotypotherium
s.p del Plioceno de la Prov. de Buenos Aires. Museo de La
Plata. |
|
Posible aspecto de Pseudotypotherium. (*). |
Mamífero
Notoungulado.
Se trata de un mamífero notoungulado perteneciente a
una antigua estirpe que evolucionó a partir del Paleoceno
sudamericano. Era un animal de tamańo mediano, ligeramente mayor que
una oveja actual, con una cola algo más larga, patas relativamente
desarrolladas y una cabeza cuya fórmula dentaria recuerda a la de
los roedores. Presentaba un cuello corto, un cuerpo robusto y, en el
extremo de sus patas, garras muy conspicuas. Su dieta era herbívora,
basada principalmente en pasturas y hojas, abundantes en la llanura
pampeana. Sus hábitos podrían compararse con los del carpincho o
capibara, siendo cursorial y semiacuático. Es característico de la
Edad Montehermosense, y en áreas comprendidas entre el sur de Mar
del Plata y el norte de Miramar se han recolectado numerosos restos
fósiles asignables a este género. Géneros y especies
relacionados: Protypotherium antiquum, Typotheriopsis
chasicoensis, Protypotherium australe, Pseudotypotherium histatum
y Mesotherium cristatum.
Promacrauchenia
chapadmalense.
Kraglievich,
1930.
|
 |
 |
 |
|
Craneo y parte del
esqueleto de Promacrauchenia, Colección Museo de
Ciencias Naturales de Miramar. |
Icnitas atribuidas a Macrauchenichnus
rector. Icnogenero de Promacrauchenia. Conicet de
Mendoza. |
Aspecto de un
Macraucheniidae (*). |
Mamífero
Litopterno. Es
un género extinto de mamíferos perteneciente a la familia
Macraucheniidae, dentro del orden Litopterna de los meridiungulados,
y de menor tamańo que Macrauchenia del Pleistoceno. Su
cuerpo recordaba al de un camélido, con una trompa corta, alcanzando
aproximadamente 1,6 metros de altura, 2,5 metros de longitud y un
peso cercano a una tonelada. Esta trompa habría funcionado como un
labio prensil, permitiéndole alimentarse, acondicionar el aire
inhalado y posiblemente permanecer parcialmente sumergido,
complementada por una lengua larga y prensil. Aunque compartía
numerosas adaptaciones morfológicas con los jiráfidos, no existía
parentesco entre ambos grupos. Los fósiles de Promacrauchenia
proceden de estratos miocenos y del Plioceno temprano de
Argentina. Los macrauquénidos evolucionaron en total aislamiento en
el continente sudamericano a partir de herbívoros primitivos. Los
promacrauquenios no estaban altamente especializados, poseían una
cola de tamańo medio, patas relativamente largas —aunque no tanto
como las de sus descendientes—, tres dedos por pata y un cerebro
bien desarrollado. Habitaban ambientes de matorral, alimentándose de
hojas, brotes, tallos, frutas, helechos y hongos, siendo capaces
incluso de consumir vegetación espinosa y coriácea. Tras su
extinción, su nicho ecológico fue ocupado por litopternos de mayor
tamańo y, en la actualidad, en parte por el guanaco. En la costa
norte del golfo San Matías, en Punta Bermeja (Río Negro), se
hallaron icnitas correspondientes al Plioceno temprano, con huellas
de hasta 4 cm de profundidad que se extienden a lo largo de 1,30
metros, atribuidas a macrauquénidos del género Promacrauchenia.
Otras especies relacionadas incluyen Epitherium laternarium,
Eoauchenia primitiva, Diplasiotherium robustum, Neolicaphrium recens,
Promacrauchenia antiqua, Promacrauchenia kraglievichi,
Promacrauchenia chapadmalense, Windhausenia delacroixi,
Pseudomacrauchenia yepesi y
Macraucheniopsis ensenadensis.
Eoauchenia primitiva.
Ameghino,
1889.
|
 |
 |
|
Algunos de los restos de un
proteroterio hallados en Termas de Río
Hondo. Museo Rincón de Atacama. |
Un grupo de
Eoauchenias
perseguidos por un ave del
terror.Por MarMag.2026 |
Mamífero
Litopterno. Se
trata de un mamífero de pequeńo tamańo, superficialmente similar a
los caballos actuales, aunque sin parentesco alguno, perteneciente a
la familia Proterotheriidae, una estirpe primitiva originaria de
América del Sur. Durante aproximadamente 50 millones de ańos de
aislamiento casi total, los mamíferos de Sudamérica y la Antártida
evolucionaron en un contexto de continente-isla, extinguiéndose en
la Antártida antes que en Sudamérica debido al enfriamiento
climático y la glaciación iniciados hace unos 37 millones de ańos.
Los proteroterios constituyen uno de los ejemplos más claros de
convergencia adaptativa o evolución paralela, al desarrollar rasgos
similares a los caballos de otros continentes pese a su aislamiento
biogeográfico. Estos pequeńos “falsos caballos” eran animales
gráciles, de lomo relativamente corto, extremidades alargadas con
pezuńas y un rostro no particularmente largo, con ojos grandes. En
conjunto, recuerdan más a gacelas u otros herbívoros pequeńos o
medianos que a caballos modernos. Las proporciones de sus miembros
sugieren que habitaban ambientes más boscosos, lo que permite
realizar inferencias paleoambientales y paleoclimáticas. Un rasgo
compartido con los caballos es la presencia de un mecanismo de
trabado en la rodilla, que les habría permitido permanecer de pie
con menor gasto energético, característica propuesta para varios
mamíferos sudamericanos extintos. En 2015 se dieron a conocer los
primeros restos de esta familia en el Mio-Plioceno de la provincia
de Santiago del Estero, procedentes de la Formación Las Cańas, en
una de las márgenes del río Dulce, cerca de la localidad de Río
Hondo, y depositados en el Museo Municipal Rincón de Atacama. Otra
localidad clásica para el hallazgo de Eoauchenia primitiva es
Monte Hermoso, en la provincia de Buenos Aires. Otros géneros y
especies conocidas son Diplasiotherium robustum y
Epitherium laternarium.
Actenomys latidens.
Ameghino, 1888.
| |
|
|
 |
|
 |
|
Ejemplar en el Museo de Ciencias Naturales de Miramar. |
|
Ilustración del extinto roedor Actenomys. Por MarMag.2025. |
Mamífero
Roedor. Fue un
roedor caviomorfo muy común en la región pampeana y considerado un
fósil guía. Se trata de un género extinguido a comienzos del
Pleistoceno, emparentado con el actual género Ctenomys
(tucos-tucos), ampliamente diversificado en Sudamérica.
Probablemente presentaba hábitos subterráneos, ya que sus restos
suelen hallarse en antiguas madrigueras. Su alimentación era
herbívora, basada principalmente en raíces y bulbos. Las
extremidades anteriores poseían falanges ungueales muy
desarrolladas, adaptadas a la excavación. El cuerpo era
relativamente corto pero ancho, con vértebras cervicales cortas y
escasa movilidad, por lo que Actenomys debía girar
completamente el cuerpo para mirar hacia atrás. Su tamańo duplicaba
ampliamente al de sus parientes actuales y, durante el Plioceno, fue
una presa frecuente de numerosos depredadores. En algunos casos, sus
dientes aparecen junto a huesos triturados dentro de excrementos
atribuidos a marsupiales y prociónidos. Su extinción estaría
asociada a la desaparición de otros taxones contemporáneos. Su
origen fue alóctono, con antecesores que habrían ingresado al
continente durante el Oligoceno medio. Se han recuperado centenares
de restos, incluidos esqueletos, cráneos y mandíbulas, en las
formaciones Chapadmalal y Vorohué, entre Mar del Plata y Miramar,
así como en otros puntos de la provincia de Buenos Aires. Géneros
relacionados: Abolosia castellanosi, Ctenomys (Paractenomys)
chapadmalensis, Eucelophorus cabrerai, Phtoramys homogenidens,
Pseudoplateomys formosus, Actenomys latidens, Ctenomys talarum
y Xenodontomys ellipticus.
Phugatherium novum.
Ameghino, 1908.
 |
 |
|
Ramas mandibulares de Phugatherium novun
del Plioceno, en el Museo de Ciencias Naturales de Miramar.,
|
Reconstrucción en vivo de Phugatherium novun. Por
MarMag.2025.: |
Mamífero
Roedor. Se
trata de una forma primitiva de los actuales carpinchos o capibaras,
aunque de mayor tamańo, con una contextura comparable a la de un
tapir asiático. Sus primeros registros corresponden al Mioceno, hace
aproximadamente 9 millones de ańos, y se extiende hasta el Plioceno,
hace unos 3 millones de ańos. El cráneo presenta un rostro alargado
y estrecho, con una longitud que duplica la del carpincho actual,
superando los 50 centímetros. El fémur y el húmero mantienen
proporciones similares a las del género moderno, pero el cúbito,
radio, tibia, peroné y otros huesos de las extremidades eran
considerablemente más largos y desarrollados, lo que le confería un
aspecto de carpincho de patas largas. Esta morfología sugiere
afinidades con mamíferos corredores y un peso que superaba los 200
kilogramos. La dentición era típica de los roedores, con incisivos
largos y desarrollados y una serie molariforme laminada, mostrando
marcadas diferencias entre individuos juveniles y adultos, lo que
durante décadas llevó a confusiones taxonómicas y a la creación de
géneros y especies hoy considerados sinónimos. Su dieta se basaba
principalmente en vegetales de zonas pantanosas, aunque algunos
paleontólogos descartan hábitos anfibios estrictos. Los restos más
importantes provienen de afloramientos pliocenos de la provincia de
Buenos Aires, como Miramar y Monte Hermoso. El carpincho actual (Hydrochoerus
hydrochaeris) no posee registros fósiles en esta región y su
presencia es muy reciente y con poblaciones reducidas. Géneros y
especies relacionadas: Anchimysops villalobosi, Anchimysops
ultra, Phugatherium cataclisticum, Protohydrochoerus perlurbidus,
Chapalmatherium novum y
Neochoerus sp.
Eumegamys paranensis. Kraglievich, 1926.
|
 |
calco%20mandibula%20mioceno%20chasico%20bs%20as.JPG) |
 |
|
Cráneo del gigantesco
Eumegamysops exhibido en el Museo de La Plata, procedente de la
localidad de Paraná, comparado con el cráneo del roedor
viviente mas grande del planeta, el carpincho. |
Carlesia sp (=eumegamys)
mandíbula del Mioceno de Chasico (Prov. de Bs As). Museo La
Plata. |
Tamańos comparativos con un
humano y un carpincho, el roedor viviente de mayor
dimensión. Por
MarMag.2025. |
Mamífero
Roedor. Eran
roedores gigantes que habitaron la región pampeana principalmente
durante el Plioceno temprano a tardío, entre hace 4 y 2 millones de
ańos, aunque también se han registrado restos aislados en otros
sectores de Sudamérica. Poseían incisivos de varios centímetros de
longitud, mientras que los molares eran relativamente pequeńos, lo
que sugiere una dieta basada en vegetales tiernos, frutos o plantas
acuáticas. En la actualidad, el mayor roedor viviente es el
carpincho, que alcanza unos 60 kilogramos, mientras que
Eumegamys superaba los 150 kilogramos. Existió durante el
Plioceno temprano a tardío, con hallazgos importantes en la
Mesopotamia argentina (Entre Ríos, Buenos Aires) y otros lugares de
Sudamérica. Este género también ha sido conocido bajo otras
denominaciones taxonómicas, tales como Megamys, Isostylomys,
Carlesia, Diaphoromys, Rusconia y
Protomegamys.
Telicomys giganteus.
Ameghino, 1904.
|
 |
|
 |
|
Mandíbulas de Telicomys en la exhibición de roedores
fósiles del Museo de Ciencias Naturales de Miramar. Copias. |
|
Posible
aspecto del gigantesco roedor Plioceno Telicomys. (*). |
Mamífero Roedor.
Es un género extinto de roedores
sudamericanos que alcanzó un tamańo extraordinario durante el
Plioceno, con especies que llegaron a dimensiones comparables a las
de una vaca doméstica. Uno de los representantes más conocidos fue
Telicomys giganteus, que poseía extremidades alargadas
y probablemente estaba adaptado a la carrera. Las mandíbulas
presentaban grandes incisivos, con una cara anterior de hasta 3
centímetros de ancho, acompańados por cuatro molares en cada maxilar
y rama mandibular. Se presume que habitaba ambientes secos con
vegetación abundante, aunque la evidencia fósil disponible es
limitada y no permite reconstrucciones biológicas detalladas.
Algunos restos significativos fueron descubiertos a mediados de la
década de 1940 en las cercanías de la ciudad de Miramar, en la
provincia de Buenos Aires. Algunos autores sugieren que pudo haber
superado los 2 metros de longitud corporal. Su pariente viviente más
cercano es la pacarana (Dinomys branickii). Géneros y
especies relacionados: Telicomys gigantissimus y
Telicomys amazonensis (especie considerada dudosa).
Paleocavia impar. Ameghino, 1889.
|
 |
|
 |
|
Cráneos de
cavidae del Plioceno. Museo de Miramar. |
|
Aspecto de
Paleocavia. (*). |
Mamífero Roedor.
Fue un género muy abundante
desde el Mioceno hasta el final del Plioceno, aunque esta familia
aún requiere estudios más profundos. Como roedores cávideos, pueden
considerarse indicadores de ambientes templado-cálidos. Presentaban
una cabeza grande y de forma rectangular, con una cola muy reducida
o ausente. Poseían cuatro dedos en las extremidades anteriores y
tres en las posteriores, un cuerpo alargado y extremidades delgadas
y cortas, que sin embargo les permitían desplazarse con notable
rapidez. Habitaban zonas con vegetación abundante y condiciones
relativamente húmedas. Alcanzaban unos 25 centímetros de longitud y
estaban emparentados con los actuales conejillos de Indias, cuises y
el carpincho, el mayor roedor viviente. Otros géneros relacionados
incluyen Microcavia, Pascualia, Dolicavia, Caviodon,
Palaeocavia, Allocavia y Galea.
Eumysops gracili.
Ameghino, 1906.
|
 |
|
 |
|
Cráneo de
Eumysops. Hallado por Francisco De Cianni. Museo de Ciencias
Naturales de Miramar. |
|
Aspecto que
presentaría Eumysops. Por MarMag.2025. |
Mamífero Roedor.
Los equimíidos son una familia
de roedores histricomorfos distribuida en Centro y Sudamérica, con
un rico registro fósil desde el Mioceno tardío en Patagonia, Mendoza
y Catamarca, y especialmente abundante en el Plioceno de Buenos
Aires y Entre Ríos. El género Eumysops incluyó una
gran diversidad de especies de octodontoideos. La mayoría de los
equimíidos se encontraba restringida a ambientes con abundante
disponibilidad de agua, mostrando escasa adaptación a regiones
áridas. Eran casi exclusivamente herbívoros; algunas especies eran
terrestres o cavadoras, mientras que otras presentaban hábitos
arbóreos. Poseían un pelaje fuerte y espinoso que actuaba como
mecanismo de defensa. Muchos equimíidos podían desprenderse de la
cola al ser atacados, lo que distraía al depredador el tiempo
suficiente para escapar, aunque dicha estructura no se regeneraba.
Otras especies conocidas son Eumysops laeviplicatus, Eumysops
formosus y Eumysops chapalmalensis.
Thylacosmilus atrox. Riggs,
1933.
|
|
|
|

|

|
|
Thylacosmilus
recreado in situ en el Museo de Miramar, por
el Técnico Mariano Magnussen Saffer. Presentado en las JAPV.
2014 |
Aspecto de Thylacosmilus.
Nótese el detalle del cachorro asomando del marsupio (*). |
Mamífero
Marsupial.
Fue un importante carnívoro marsupial perteneciente
a los Sparassodonta de la Era Terciaria, muy similar en aspecto al
denominado “tigre dientes de sable”, aunque sin parentesco alguno,
ya que este último es un mamífero placentario. Constituye un claro
ejemplo de evolución paralela: dos linajes totalmente distintos que
nunca coexistieron en el mismo continente desarrollaron morfologías
semejantes al ocupar un rol ecológico equivalente.
Thylacosmilus poseía largos y afilados colmillos,
proyectados hacia abajo y hacia adentro, de aproximadamente 15
centímetros de longitud, y un tamańo corporal algo menor al de un
puma actual. Los primeros restos fósiles de este impresionante
carnívoro fueron hallados en la década de 1920 en áreas de Entre
Ríos, así como en el valle de Santa María y Puerta de Corral
Quemado, en el departamento de Belén, provincia de Catamarca. A
diferencia de Smilodon, Thylacosmilus
carecía de incisivos y de una vaina protectora para los caninos, los
cuales crecían de manera continua, como los dientes de los roedores.
Es muy probable que sus presas fueran grandes mamíferos
notoungulados, a los que abatía mediante una mordida certera en el
cuello. Sus caninos habrían provocado heridas profundas que
ocasionaban una rápida hemorragia, llevando a la muerte del animal
en pocos segundos. En Sudamérica se han recuperado escasos restos,
procedentes de Entre Ríos, Córdoba y Catamarca; el cráneo más
completo proviene del Plioceno de Chapadmalal, y se conocen restos
de un esqueleto parcial del Plioceno de Miramar. Su extinción se
asocia a los profundos cambios ambientales ocurridos hacia el final
del Plioceno y a la desaparición de las grandes presas que acechaba.
Thylacosmilus atrox habría tenido una masa corporal estimada
entre 90 y 140 kilogramos. Géneros relacionados: Thylacosmilus
lentis y Achlysictis lelongi.
Thylatheridium cristatum.
Reig, 1952.
|
|
|
|
|
 |
|
 |
|
Mandíbula de Thylatheridium. Museo de Ciencias Naturales
de Miramar "Punta Hermengo". |
|
Aspecto de
Thylatheridium cristatum.
Por MarMag.2025. |
Mamífero Marsupial. Fue un animal
perteneciente a la familia de los marsupiales didélfidos. Su cráneo
era relativamente grácil y pequeńo, con una caja cerebral estrecha,
y un hocico puntiagudo. La cola era larga y prensil, como en otros
didélfidos actuales. Este grupo presentaba una notable capacidad de
adaptación a distintos ambientes, aunque probablemente prefería
zonas arboladas cercanas a fuentes de agua. Necesitaba disponer de
refugios seguros que le brindaran protección durante el día y abrigo
en invierno, estación especialmente crítica, ya que no toleraba bien
las bajas temperaturas. Sin embargo, no excavaba madrigueras
propias, sino que utilizaba refugios naturales, como cuevas en rocas
o troncos de árboles, o bien ocupaba refugios abandonados por otros
animales. El registro fósil sugiere que Thylatheridium
tenía hábitos mayormente solitarios. Era omnívoro y poseía una
dieta muy variada, lo que le permitía adaptarse a distintos hábitats.
Consumía frutos maduros, brotes y tallos tiernos, así como
invertebrados, insectos, lombrices y, con frecuencia, pequeńas aves,
especialmente durante la época reproductiva. Se han recuperado
restos bien preservados de este género en los afloramientos del
litoral marítimo bonaerense, en Barranca de los Lobos, Chapadmalal y
Miramar, así como en sedimentos terciarios del Quequén Salado y
Monte Hermoso. Géneros relacionados: Didelphis albiventris,
Didelphis crucialis, Didelphis reigi, Hyperdidelphys inexpectata,
Lestodelphys halli, Lestodelphys juga, Lutreolina crassicaudata,
Lutreolina tracheia, Monodelphis dimidiata, Thylamys pusillus,
Thylamys contrerasi, Thylatheridium pascuali, Thylophorops perplata
y Thylophorops chapadmalensis.
Argyrolagus palmeri.
Ameghino, 1904.
 |
 |
 |
|
Cráneo de
Argyrolagus de la
Col. del Museo de La Plata. |
Esqueleto
de Argyrolagus recreado a partir de los fósiles
conocidos. Museo de Ciencias Naturales de Miramar. |
Aspecto de
Argyrolagus.
Ambas recreaciones de Mariano Magnussen.
Por MarMag.2026 |
Mamífero Marsupial. Fue
un mamífero marsupial de pequeńo tamańo, comparable al de un ratón
grande, con una longitud aproximada de 40 centímetros, emparentado con
los actuales canguros de Australia y las zarigüeyas de América del Sur.
Su aspecto recuerda notablemente al de las ratas canguro modernas.
Poseía largas patas traseras y extremidades anteriores cortas, con
mandíbulas de aspecto roedoriforme. Es posible que tuviera orejas
largas, aunque estas no se han preservado en el registro fósil. Se
alimentaba de vegetales e insectos y probablemente tenía hábitos
crepusculares o nocturnos. Argyrolagus formó parte de una
notable radiación evolutiva independiente que tuvo lugar en Sudamérica
durante los últimos 60 millones de ańos, cuando el continente permaneció
aislado del resto de las masas continentales y desarrolló una fauna
única. Estos marsupiales ocuparon nichos ecológicos que en otros lugares
del mundo estaban dominados por mamíferos placentarios. El primer
hallazgo de este género se realizó en el yacimiento de Monte Hermoso, al
sur de la provincia de Buenos Aires. Resulta llamativo que el nicho
ecológico ocupado por Argyrolagus no haya sido reemplazado
por otra especie tras su desaparición. Géneros relacionados:
Argyrolagus palmeri (Edad Montehermosense), Argyrolagus
scagliari (Edad Chapadmalalense) y Microtragulus reigi
(formaciones Chapadmalal, Vorohué, Barranca de los Lobos y San Andrés).
También se han recuperado restos en sedimentos pliocenos de Miramar,
Quequén Grande y Pehuen Co.
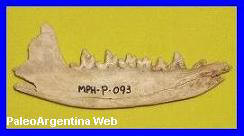
Thylophorops chapadmalensis.
Ameghino, 1908.
 |
 |
 |
|
Coprolitos y otros desechos
escatológicos atribuido a
Thylophorops. Plioceno de Miramar. |
Rama mandibular de
Thylophorops. Colección Museo de Ciencias Naturales de
Miramar. |
Posible aspecto de
Thylophorops. Por MarMag.2025. |
Mamífero Marsupial. Es un
género extinto de marsupiales didelfimorfos de la familia
Didelphidae, estrechamente emparentado con las zarigüeyas modernas
de los géneros Philander y Didelphis.
Sus fósiles proceden de la provincia de Buenos Aires, Argentina. Una
de sus especies, Thylophorops lorenzinii, es el mayor
didélfido conocido, con un peso estimado entre 4,7 y 7,5 kilogramos,
superando al actual Didelphis virginiana. Un hallazgo
publicado recientemente sugiere que un ejemplar de
Thylophorops chapadmalensis habría sido depredado por
Cyonasua lutaria, ya que ambos fueron encontrados asociados
a otras especies dentro de una cueva o crotovina atribuida a un gran
armadillo. A este género también se le atribuyen varios coprolitos
fósiles hallados en el interior de dicha estructura.
Hyperdidelphys inexpectata.
Ameghino, 1889.
|
 |
|
 |
|
Mandíbula
inferior de Hyperdidelphys del Plioceno de Miramar. |
|
Aspecto de
Hyperdidelphys junto a sus crías. Por MarMag.2025. |
Mamífero Marsupial. Es
un género extinto de marsupiales didelfimorfos de la familia
Didelphidae cuyos fósiles, datados entre el Mioceno Superior y el
Plioceno Superior, proceden del centro y noreste de Argentina. Las
adaptaciones carnívoras de las especies de Hyperdidelphys
se vinculan con el proceso de declinación de los Sparassodonta
durante el Mioceno tardío y el Plioceno en América del Sur, previo a
la llegada de los carnívoros placentarios de origen holártico tras
la formación del istmo de Panamá. Probablemente acechaba y atacaba
dentro de madrigueras, depredando pequeńos roedores, armadillos,
notoungulados y anfibios, entre otros vertebrados.
Lutreolina tracheia.
Rovereto 1914.
|
 |
|
.jpg) |
|
Restos recreados in situ
del genero Lutreolina sp. de Ciencias Naturales de Miramar. |
|
Aspecto general del genero
Lutreolina sp del Mioceno - Holoceno.
Por Carlos C. Wiedner.
|
Mamífero Marsupial. Es un género
de mamíferos marsupiales conocido en América del Sur desde el
Terciario. Los registros más antiguos corresponden al Mioceno tardío
(Edad Huayqueriense) del noroeste y centro de Argentina, mientras
que Lutreolina tracheia se conoce en niveles pliocenos
de las edades Montehermosense y Chapadmalalense en las provincias de
Buenos Aires, Córdoba y Catamarca. Al igual que sus parientes
actuales, probablemente habitaba pastizales asociados a humedales de
tierras bajas. Presentaba una cabeza pequeńa, patas cortas y un
cuerpo alargado. Su dieta incluía pequeńos mamíferos, aves y sus
nidos, anfibios, reptiles, peces, moluscos e insectos, complementada
en menor medida con alimentos de origen vegetal. El género también
está representado durante el Pleistoceno y el Holoceno. En la
actualidad se reconocen dos especies vivientes: Lutreolina
crassicaudata y Lutreolina massoia.
Chapalmalania altaefrontis.
Ameghino,1908.
|
|
|
|
 |

|
|
Cráneo de
Chapadmalania superponiendo las marcas dentales en
hueso de
Eosclerocalyptus. Tomado de Paleo Electrónica. |
Reconstrucción
de Chapadmalania. Por MarMag.2025. |
Mamífero
Carnívoro.
Fue un procionido que llegó a América
del Sur durante el Gran Intercambio Biótico Americano, ocurrido en
el Plioceno, aunque su registro se extiende desde fines del Mioceno
hasta comienzos del Pleistoceno, entre 6,8 y 1,8 millones de ańos.
Se adaptó rápidamente a las nuevas condiciones ambientales,
diversificándose ampliamente en el continente. Chapalmalania
altaefrontis debió asemejarse a un mapache gigante,
alcanzando cerca de 2 metros de longitud, 1,5 metros de altura y un
peso aproximado de 160 kilogramos. Su tamańo era tal que, en los
primeros hallazgos, fue interpretado erróneamente como un oso
prehistórico. Su dieta habría sido variada, incluyendo plantas,
frutos, huevos, peces, insectos y carrońa. El cráneo era ancho y
recordaba al de un lobo, con una dentición completa: incisivos
curvados y robustos, caninos cortos y fuertes, y molares con amplias
superficies de trituración. Su similitud con el panda gigante
constituye otro ejemplo de convergencia evolutiva. Este gran
carnívoro desaparece del registro fósil hace unos 3 millones de ańos
y se conocen muy pocos restos diagnósticos. En 2014 se dio a conocer
el hallazgo de restos fósiles en Olavarría que evidencian la
depredación de un Eosclerocalyptus lineatus por
Chapalmalania.
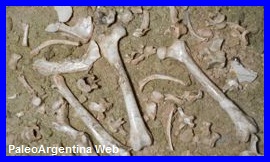
Cyonasua lutaria.
Cabrera,
1936.
 |
 |
 |
|
Esqueleto parcialmente completo de Cyonasua
lutaria. Colección del Museo de Ciencias
Naturales de Miramar.. |
Cráneo de
Cyonasua, en las colecciones del Muso de La
Plata. |
Reconstrucción en vivo del
prociónido
Cyonasua del Plioceno bonaerense. Por
MarMag.2025. |
Mamífero Carnívoro.
Es una especie extinta de
carnívoro sudamericano cuyo nombre, de origen griego,
significa “coatí-perro”. Fue un procionido, al igual que
Chapalmalania, aunque de menor tamańo, y
estaba emparentado con el actual género Nasua,
presente en la selva amazónica y misionera. Formó parte
de la fauna alóctona cuya estirpe evolucionó en
Sudamérica desde el Mioceno, hace aproximadamente 10
millones de ańos, siendo uno de los primeros carnívoros
placentarios en colonizar el continente. Sus restos han
sido hallados en sedimentos de las provincias de
Catamarca, Mendoza, Córdoba, Buenos Aires y La Pampa. A
diferencia de otros carnívoros, Cyonasua lutaria
poseía caninos muy desarrollados y robustos, y
probablemente se alimentaba de pequeńos mamíferos, como
Paedotherium o los antecesores de los
armadillos actuales, aunque también consumía frutos,
huevos y larvas. Sus primeros fósiles fueron descritos
en Argentina en 1885, y la especie habría vivido entre
3,6 y 2,5 millones de ańos. Fue un componente relevante
del Gran Intercambio Biótico Americano, ya sea mediante
el puente terrestre del istmo de Panamá o a través de
dispersión insular. En Miramar se ha recuperado parte de
un esqueleto, actualmente preservado en el Museo de
Ciencias Naturales de esa localidad.
Conepatus altiramus.
Reig, 1952.
|
 |
|
 |
|
Recreación in
situ del genero Conepatus. Museo de Ciencias
Naturales de
Miramar. |
|
Posible
aspecto del genero Conepatus del Plio- Pleistoceno de la
región pampeana (*). |
Mamífero
Carnívoro. Los zorrinos de América
del Sur, tanto fósiles como actuales, son asignados al género
Conepatus. La familia Mephitidae agrupa carnívoros pequeńos a
medianos, de cuerpo alargado, patas cortas y cola bien desarrollada,
caracterizados por la presencia de glándulas anales que producen
secreciones de olor intenso. En sedimentos referibles al Plio-Pleistoceno
temprano y medio de las barrancas de la costa atlántica, cerca de
Miramar, se ha registrado la especie Conepatus altiramus,
considerada posiblemente ancestral de los zorrinos sudamericanos
actuales. Antiguamente, los mefítidos eran incluidos dentro de la
familia Mustelidae, pero estudios filogenéticos recientes
demostraron que Mephitidae constituye un grupo hermano de Mustelidae
y Procyonidae, motivo por el cual fue elevada al rango de familia
independiente. Los registros más antiguos de mefítidos provienen del
Ensenadense (Pleistoceno temprano-medio), y restos fósiles de
Conepatus se conocen en Venezuela, Perú, Bolivia, Brasil y
Argentina.
Pyramiodontherium bergi.
Moreno & Mercerat, 1891.
 |
 |
 |
|
Femur de
Pyramiodontherium
sp. Museo Paleontologico de San Pedro. |
Huellas de Pyramiodontherium sp,
atribuidas al icnofosil
Megatherichnum oportoi, Mioceno de Carmen de
Patagones (provincia de Buenos Aires). |
Posible aspecto del perezoso
gigante del Mio-Plioceno
Pyramiodontherium
sp. (*): |
Mamífero Xenarthro. Se trata de un
xenartro emparentado con los Megatheriinae, frecuentes
representantes fósiles en los afloramientos geológicos del Mioceno
tardío–Plioceno temprano de las provincias de Catamarca y Tucumán.
Restos muy completos y de alto valor paleontológico fueron
rescatados recientemente en niveles informalmente denominados “Araucarense”,
en el Bajo de Andalhuala, Catamarca. Su cráneo presenta un rostro
más ancho que largo. Este interesante perezoso extinto poseía
falanges ungueales muy desarrolladas, al igual que otras especies
extintas y vivientes. Su dieta habría estado constituida por la
vegetación abundante en la región durante el Terciario, aunque
también podría haber incluido carrońa, tal como se ha demostrado en
otros representantes del infraorden Pilosa. El fémur era corto y
robusto, recordando fácilmente a los gigantes del Pleistoceno. En
esta región también vivió otro gran taxón emparentado con el género
en cuestión, denominado Plesiomegatherium hansmeyeri,
y Megatherium sp. fue un típico megamamífero del
noroeste argentino. En el Mioceno tardío, en el norte de la
Patagonia, cerca de Carmen de Patagones (provincia de Buenos Aires),
se han registrado numerosas huellas de Megatherichnum oportoi
Casamiquela, 1974, atribuidas al género
Pyramiodontherium sp.
Proscelidodon patrius.
Ameghino, 1888.
|
 |
|
 |
|
Cráneo de
Proscelidodon patrius del Plioceno argentino, Col. Museo
Municipal de Miramar. |
|
Reconstrucción de Proscelidodon, por el paleoartista
Jorge Blanco. |
Mamífero Xenarthro.
Es tal vez el xenartro más característico de los
sedimentos del Terciario superior de la región pampeana, con un
parentesco cercano a Scelidotherium y Scelidodon
del Pleistoceno. Se trata de un enorme Pilosa que habitó estas llanuras
durante el Plioceno tardío. Se caracterizaba por un cráneo macizo y
alargado, un cuerpo rechoncho y de aspecto robusto, y extremidades
cortas y poderosas. Probablemente estaba cubierto por una larga
pelambre, como en otros perezosos mejor conocidos. Su alimentación
habría incluido pastizales extensos, vegetación palustre y,
posiblemente, carrońa. Es probable que pudiera erguirse sobre las patas
traseras y también excavar largas galerías subterráneas para refugiarse.
Sus restos son hallados con frecuencia en las cercanías de las
localidades fosilíferas de Chapadmalal y Monte Hermoso, en la provincia
de Buenos Aires. Géneros relacionados incluyen Proscelidodon
patrius, Scelidotherium floweri, Scelidotherium parodi, Scelidodon
patrius, Scelidotherium leptocephalum y
Scelidodon capellini.
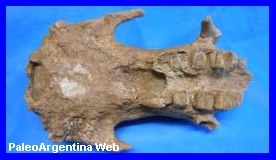
Megatherium sp.
Cuvier, 1796.
|
 |
|
 |
|
Cráneo de Juvenil de
Megatherium sp del Plioceno.
Museo de Cs Naturales
de Miramar. Es el mas antiguo de Argentina. |
|
Reconstrucción de Megatherium sp del Plioceno.
(*). |
Mamífero Xenarthro.
Se trata de otro Pilosa de gran tamańo, que alcanzaba
aproximadamente 4,5 metros de longitud y un peso cercano a los 500
kilogramos. Su hábitat habría estado conformado por bosques de
algarrobos y quebrachales, bajo un clima más cálido y húmedo que el
actual. Carecía de enemigos naturales importantes, ya que los
principales carnívoros de la época eran marsupiales de tamańo similar al
de un perro grande. Posiblemente se trataba de mamíferos poco sociables
y de hábitos mayormente vegetarianos. Eran animales netamente
terrestres, ya que su gran tamańo les impedía llevar un modo de vida
arborícola como el de sus parientes actuales. Se alimentaban en posición
bípeda. Este Megatherium presentaba un rostro más ancho
que largo, y los molares superiores tenían una sección cuadrangular. Los
perezosos terrestres, como otros xenartros, evolucionaron en aislamiento
en Sudamérica, que durante el Paleógeno funcionó como un
continente-isla. Durante el Plioceno, la formación del istmo de Panamá
desencadenó el Gran Intercambio Biótico Americano y una extinción masiva
de buena parte de la megafauna autóctona. Sin embargo, los perezosos
terrestres se vieron relativamente poco afectados y continuaron
prosperando, incluso migrando hacia Norteamérica, donde subsistieron
hasta fines del Pleistoceno. El reciente hallazgo de Megatherium
sp. del Plioceno, dado a conocer por investigadores del CONICET,
Fundación Azara, LACEV y el Museo de Ciencias Naturales de Miramar,
corresponde a los restos fósiles más antiguos conocidos para este
género, junto con materiales de Bolivia y Perú. El cráneo pertenece a un
ejemplar juvenil, lo que impidió determinar si se trata de una nueva
especie. Con el tiempo, las especies de megaterios aumentaron
progresivamente su tamańo, alcanzando su máximo desarrollo en el
Pleistoceno tardío con Megatherium americanum, que llegó a
dimensiones comparables a las de un elefante africano. Sus poderosas
mandíbulas poseían 16 molares carentes de esmalte. Es posible que las
formas pliocenas también recurrieran a la carrońa, como otros perezosos
gigantes extintos.
Myrmecophaga carolaameghinoi. Kraglievich, 1934.
|
 |
|
 |
|
Esqueleto de
Myrmecophaga tridáctila, en el
Museo Argentino de Ciencias Naturales. Ilustrativo. |
|
Aspecto de
Myrmecophaga. Por MarMag.2025. |
Mamífero Xenarthro.
Se trata de un integrante extinto de la familia
Myrmecophagidae, correspondiente al Plio-Pleistoceno bonaerense. Se
lo conoce a partir del hallazgo de un metacarpiano procedente de la
localidad de Monte Hermoso, de tamańo algo menor al de los
ejemplares actuales. Por analogía con su representante viviente,
Myrmecophaga tridactyla, Myrmecophaga
caroloameghinoi habría poseído un hocico alargado y convexo,
especialmente adaptado para su tipo de alimentación. Presentaba una
cola prensil, peluda en la base y desnuda en el extremo, cuatro
garras largas y poderosas en las extremidades anteriores y cinco más
pequeńas en las posteriores. Un ejemplar adulto pudo haber alcanzado
1,5 metros de longitud total, con una lengua de hasta 50
centímetros. Su visión habría sido débil, compensada por sentidos
más desarrollados, aunque estas inferencias son hipotéticas debido a
la escasez de restos. Carecía de dientes y se alimentaba de hormigas
y termitas. Sus parientes filogenéticos más cercanos son los
armadillos y los perezosos arborícolas. La presencia de un oso
hormiguero en el sur de la provincia de Buenos Aires indica
condiciones climáticas muy distintas a las actuales, probablemente
templadas a cálidas, con ambientes abiertos similares a los del
Chaco paraguayo. Esta especie fue descripta originalmente en 1934
por Lucas Kraglievich bajo el nombre Nunezia caroloameghinoi,
siendo posteriormente reasignada al género Myrmecophaga
en 1976 por Sue E. Hirschfeld, quien destacó su menor tamańo
respecto de la especie viviente.
Ringueletia simpsoni.
Bordas, 1933.
|
 |
|
 |
|
Conjunto de restos fósiles
de Ringueletia simpsoni, del Plioceno de Miramar.
Museo Punta Hermengo. |
|
Posible aspecto de
Ringueletia simpsoni. asechado por
Thylacosmilus en el Plioceno bonaerense. |
Mamífero Xenarthro.
Fue un armadillo de gran
tamańo cuyos restos, recuperados recientemente en el Plioceno de la
ciudad de Miramar y asociados a otros vertebrados contemporáneos,
constituyen la primera evidencia directa que vincula a estos grandes
armadillos extintos con paleocuevas o crotovinas de esta edad
geológica. Su masa corporal fue estimada en aproximadamente 40
kilogramos. Aunque su paleobiología aún no ha sido estudiada en
profundidad, el marcado desarrollo de las crestas de inserción
muscular del húmero, junto con la presencia de falanges ungueales
carpales comprimidas y acuminadas, sugiere un marcado hábito
cavador. En este contexto, el conjunto de evidencias disponibles
permite postular a Ringueletia simpsoni como el
constructor de grandes galerías subterráneas, utilizadas tanto como
refugio como para la protección de su descendencia, aunque en
algunos casos pudieron convertirse en trampas mortales. Presentaba
una dieta variada. Entre las especies relacionadas se incluyen
Holmesina paulacoutoi, Kraglievichia paranensis, Scirrotherium,
Vassallia y Plaina.
Paraeuphractus
sabateri.
Esteban, G.
2006.
|
Ejemplares de
Paraeuphractus sabateri
en el Museo Municipal Rincón de Atacama, Termas de Rió
Hondo, provincia de Santiago del Estero, e ilustración de su
posible aspecto.
Por MarMag.2025. |
Mamífero Xenarthro.
Es un mamífero cingulado
representado por los quirquinchos o armadillos actuales de gran
parte de Sudamérica, un grupo muy característico de la fauna
neotropical. Se distinguen por poseer un caparazón dorsal formado
por placas yuxtapuestas, generalmente dispuestas en filas
transversales, una cola relativamente larga y extremidades cortas.
El nuevo taxón amplía la distribución geográfica del género más allá
de los valles de altura de Catamarca y Tucumán y es considerado una
especie montehermosense del Plioceno temprano. Paraeuphractus
sabateri procede de la Formación Las Cańas, en las
proximidades de la ciudad de Las Termas de Río Hondo, provincia de
Santiago del Estero. Se recuperaron tres ejemplares, dos de ellos
con el caparazón dorsal casi completo, además de dos cráneos, una
hemimandíbula y diversos restos postcraneanos. El epíteto específico
honra a su descubridor, Sebastián Sabater, director del Museo de
Termas de Río Hondo. Su alimentación habría sido variada, como en
otros armadillos. Entre las especies relacionadas se incluyen
Paraeuphractus prominens, Doellotatus chapalmalensis,
Macrochorobates chapalmalensis, Chorobates recens, Tolypeutes sp.,
Chaetophractus villosus y
Zaedyus pichiy.
|
 |
 |
 |
|
Cráneo de
Macroeuphractus
s.p (*). |
Fragmento de
coraza de
Macroeuphractus s.p
del Mio-Plioceno de Termas de Rió Hondo.
|
Aspecto del armadillo depredador
Macroeuphractus.
|
Mamífero Xenarthro.
Es un género de armadillos
extintos registrado desde el Mioceno tardío hasta el Plioceno tardío
de Sudamérica. Se caracteriza por su gran tamańo, ya que
Macroeuphractus outesi es el armadillo no pampatérido ni
gliptodonte más grande conocido. Una de sus principales
especializaciones es su dieta carnívora, única entre los armadillos.
Actualmente se reconocen tres especies: M. outesi, M. retusus
y M. moreni. La especie tipo, M. outesi,
se conoce a partir de un espécimen del Plioceno tardío de la
provincia de Buenos Aires, compuesto por un cráneo bien conservado y
numerosos elementos del esqueleto, con un peso estimado cercano a
los 100 kilogramos. Macroeuphractus retusus se conoce
únicamente por una mandíbula inferior de la Formación Cerro Azul, en
el centro de Argentina, mientras que M. moreni ha sido registrada en
diversas formaciones del Mioceno tardío y Plioceno de Argentina y
Bolivia. Como la mayoría de los armadillos, era fosorial y
probablemente excavaba para capturar mamíferos pequeńos y medianos,
como roedores caviomorfos, pequeńos notoungulados y argirolagoideos.
Debido a su tamańo, especies como M. outesi habrían
ocupado un rol de depredador dentro de sus comunidades faunísticas,
tal como seńalaron Sergio F. Vizcaíno y Gerardo De Iuliis.
Chorobates recens.
Ameghino
1887.
|
 |
 |
 |
|
Cráneo y coraza cefalica de Chorobates recens
del Plioceno bonaerense. Museo de La Plata. |
Fragmento de
coraza de
Chorobates
s.p
del Mio-Plioceno de Termas de Rió Hondo.
Santiago del Estero. |
Aspecto de
Chorobates recens.
Por MarMag.2025. |
Mamífero Xenarthro.
En 1887, Florentino
Ameghino reconoció una nueva especie de Dasypodidae, a la que
denominó Proeuphractus recens, basándose en placas del
caparazón dorsal provenientes de sedimentos pliocenos de Monte
Hermoso, provincia de Buenos Aires. Construía amplias madrigueras
que eran reutilizadas por otros mamíferos y reptiles, encontrándose
en algunos casos restos fósiles de estos ocupantes en su interior.
Su alimentación habría sido variada, como en otros armadillos,
siendo probablemente insectívoro y omnívoro, con hábitos necrófagos
característicos de ambientes tropicales y subtropicales de
Sudamérica.
Chlamydotherium paranense.
Ameghino, 1883.
| |
 |
 |
 |
|
Fragmento de cráneo de
Pampatheriidae
Houston Museum of Natural History. Ilustrativo.
|
Pata delantera de
Pampatheriidae.
Houston Museum of Natural History. Ilustrativo. |
Aspecto de
Chlamydotherium paranense,
MarMag.2025. |
Mamífero Xenarthro.
Fue un pampatérido de gran tamańo, algo
menor que Pampatherium del Pleistoceno. Presentaba un
caparazón con escudetes escapular y pelviano bien diferenciados,
siendo el anterior corto y el posterior más prolongado, separados
por pocas bandas móviles. Estaba compuesto por placas grandes con
una figura central amplia, típica de la familia. Su cráneo era
proporcionalmente más robusto que el de Pampatherium y
poseía 36 dientes. Tenía extremidades relativamente largas en
comparación con otros dasipódidos, aunque igualmente anchas y
robustas, y una cola protegida por placas y escamas óseas y córneas.
Su dieta era muy variada, desde vegetales hasta carrońa en distintos
estados de descomposición. Entre Miramar y Mar del Plata se observa
una notable abundancia de paleocuevas atribuidas a este género o a
formas morfológicamente similares. Desde el siglo pasado, diversos
investigadores han debatido su relación filogenética, planteando si
los pampatéridos están más próximos a los gliptodontes que a los
dasipódidos. Entre los géneros relacionados se incluyen
Holmesina, Kraglievichia, Scirrotherium, Vassallia y
Plaina.
Vassallia máxima.
Castellanos, 1946.
|
 |
 |
 |
| Placa u
osteodermo de Vassallia maxima del Plioceno de
Miramar. Museo de Ciencias Naturales de Miramar. |
Cráneo de
Vassallia maxima (*). |
Posible
aspecto del enorme armadillo Vassallia maxima
tomado de Prehistoric Fauna. |
Mamífero Xenarthro.
Los Pampatheriidae
forman parte de los Xenarthra (Cingulata) nativos de América del
Sur. Tradicionalmente se los consideró estrechamente emparentados
con los armadillos actuales, aunque existe evidencia que sugiere una
relación más cercana con los gliptodóntidos. Evolucionaron en
Sudamérica durante su prolongado aislamiento cenozoico, y géneros
como Holmesina y
Pampatherium colonizaron
Norteamérica tras la formación del istmo de Panamá. Finalmente, los
pampatéridos se extinguieron en ambos subcontinentes durante el
Pleistoceno. Vassallia maxima
fue un representante plioceno registrado en las formaciones
Chiquimil y Andalhuala, en Catamarca, y constituyó un importante
herbívoro, posiblemente carrońero, con el cuerpo, la cola y el
cráneo protegidos por un caparazón. Poseía grandes garras para
excavar madrigueras, cuyas paleocuevas aún pueden reconocerse. Otros
pampatéridos del Mio-Plioceno incluyen Scirrotherium,
Chlamydotherium paranense, Kraglievichia paranensis, Plaina
y Vassallia,
esta última con dos especies reconocidas. En el Pleistoceno, el
máximo exponente de la familia fue Pampatherium typum.
Paraglyptodon chapadmalensis.
Ameghino y Rovereto,
1914.
|
 |
|

|
|
Fragmento de
rama mandibular atribuido tentativamente a
Paraglyptodon. Museo de Miramar. |
|
Reconstrucción
en vivo de Paraglyptodon chapadmalensis, común en
estratos Pliocenos de la región pampeana. MarMag.2025. |
Mamífero Xenarthro.
Es un gliptodonte cuya familia está mejor representada durante el
Pleistoceno. Fue una especie típica de la antigua Pampasia y de
tamańo considerablemente menor que Glyptodon. Se
caracterizaba por una coraza formada por placas óseas y córneas de
origen epidérmico, con un escudete cefálico que protegía la cabeza.
Su cráneo era pequeńo en relación con el cuerpo, con ramas
mandibulares muy desarrolladas y una compleja hilera de dientes sin
esmalte. La cola estaba compuesta por anillos óseos y podía
utilizarse como defensa frente a carnívoros marsupiales de la época,
como Thylacosmilus. Alcanzaba unos dos metros de
longitud y un peso aproximado de 350 kilogramos, alimentándose
principalmente de vegetales duros. Sus restos fósiles suelen ser
fragmentarios y aislados. Se extinguió hacia fines del Plioceno,
hace unos tres millones de ańos, siendo reemplazado por especies más
exitosas que se diversificaron durante el Pleistoceno.
Plohophorus figuratus.
Ameghino, 1887.
|
 |
 |
 |
|
Tubo caudal de
Plohophorus
figuratus procedente de la localidad de Monte hermoso. Museo
de La Plata. |
Fragmento de coraza de Hoplophorus euphractus, con
detalles de las placas dorsales. Genero similar. |
Imagen
ilustrativa. Publicada por Derf. |
Mamífero Cingulata.
Fue
un gliptodonte de aproximadamente dos metros de longitud, un metro
de altura y un peso cercano a los 300 kilogramos. Como otros
gliptodontes, tenía una dieta herbívora y hábitos pastoreadores,
desplazándose en ambientes abiertos dominados por vegetación
herbácea. Se destacaba por su caparazón macizo y rígido, un escudo
cefálico del mismo material y una cola robusta. Durante el
Cuaternario, algunos gliptodontes migraron hacia el norte,
alcanzando regiones centrales de los actuales Estados Unidos. Estos
animales se extinguieron hace menos de dos millones de ańos, junto
con gran parte de la megafauna. En la Edad Chapadmalalense, los
Plohophorini están representados por Plohophorus figuratus
y Plohophorus conterminus, mientras que en el
Montehermosense se distinguen Plohophorus ameghinoi y
Plohophoroides castellanos. Otra especie relacionada es
Plohophorus paranaensis.
Eosclerocalyptus lineatus.
Ameghino, 1888.
|
 |
 |
 |
|
Coraza de
Eosclerocalyptus lineatus, del
Plioceno de Olavarria. Tomado de Paleontología Electrónica. |
Coraza y
restos varios de un posible
Eosclerocalyptus sp, del Plioceno
Temprano. Museo Rincón de Atacama. |
El Neosclerocalyptus del Pleistoceno, era
relativamente semejante a su representante Plioceno.
Ilustración de
Carlos Montefusco. |
Mamífero Cingulata. Son
uno de los mamíferos herbívoros cenozoicos más reconocidos de
América del Sur, aunque durante el Plioceno eran relativamente
escasos, y llegaron a América del Norte durante el Gran Intercambio
Biótico Americano. En 2013, un equipo de paleontólogos argentinos
dio a conocer el hallazgo de una coraza y diversos elementos del
esqueleto de Eosclerocalyptus lineatus cerca de
Olavarría, en la provincia de Buenos Aires, en sedimentos poco
conocidos denominados “Cantera Avellaneda”. Lo más notable del
hallazgo fue la presencia de marcas de mordedura en varios restos,
compatibles con la dentición del procionido plioceno Chapalmalania.
Durante gran parte del Cenozoico, los carnívoros placentarios
estuvieron ausentes en Sudamérica, siendo su nicho ocupado por
marsupiales y aves corredoras. Este descubrimiento permitió
reconstruir aspectos clave de la muerte de este gliptodonte y su
ambiente. La evidencia de depredación sobre grandes mamíferos
acorazados es extremadamente escasa, limitada a pocos registros del
Plioceno y Pleistoceno, y a casos de consumo humano tardío.
Eleutherocercus sp.
Koken 1888.
 |
 |
 |
|
Mazo del tubo caudal de
Eleutherocercus setifer,
en el Museum für Naturkunde, Berlin. |
Posible cráneo de
Eleutherocercus sp, in situ. recuperado por el Museo de
Ciencias Naturales de
Miramar. |
Aspecto de un
Doedicurino como
Eleutherocercus
por
Peter Schouten. |
Mamífero Cingulata. Este
animal, relacionado con los armadillos actuales, estaba cubierto por
un caparazón y poseía un gran mazo caudal utilizado como defensa
frente a depredadores como Thylacosmilus o
Chapalmalania. Se cree que estos mazos, posiblemente más
desarrollados en los machos, también habrían intervenido en disputas
territoriales o reproductivas, de forma análoga a lo observado en el
anquilosaurio, un dinosaurio acorazado del Cretácico. Alcanzaba
cerca de 1,5 metros de altura y unos 3 metros de longitud. Era
herbívoro y habitaba bosques y praderas, con una visión
probablemente deficiente. Los gliptodontes constituyen un clado de
Cingulata caracterizado por una coraza dorsal rígida formada por
osteodermos, y actualmente se los incluye dentro de la familia
Chlamyaphoridae. Eleutherocercus fue descripto
originalmente a partir de un tubo caudal del Plioceno de Uruguay, al
que luego se sumaron caparazones parciales y osteodermos aislados.
Sus placas presentan una superficie externa lisa con grandes
perforaciones, similares a las de Doedicurus, aunque
de menor tamańo. Una cavidad esférica en la región pélvica del
caparazón dorsal probablemente alojaba una glándula principal, una
estructura casi única entre los Chlamyaphoridae. Vivió
principalmente entre el Plioceno tardío y el Pleistoceno temprano de
Uruguay y Argentina.
Platygonus
chapadmalensis.
Castellanos,
1927.
 |
|
 |
|
Cráneo y
mandíbulas de un representante de los
tayasúidos
en el Museo de Cs Naturales de Miramar. |
|
Aspecto
agresivo de un Platygonys. Por MarMag.2025. |
Mamífero Artiodactyla. Es
un género extinto de pecaríes herbívoros de la familia de los
tayasúidos, que fue un endémico de Norteamérica del periodo Mioceno
al Pleistoceno, ingresando durante el Plioceno medio a Sudamérica,
conquistando ambientes abiertos y de pastizales. Algunas especies
del Plioceno, superaban los 65 kilogramos de peso. Platygonus era
un animal gregario y, como los modernos pecaríes, posiblemente se
movía en grupos. Se extendió desde el sur de Canadá a México y de
California a Pensilvania, y llegando a las pampas de Argentina. Platygonus era
mayor que los actuales pecaríes, con cerca de 1 metro de longitud
corporal, y poseía largas patas, permitiéndole correr rápidamente.
También tenía un hocico similar al de un cerdo y largos colmillos
que probablemente usaba para defenderse de los depredadores. La
familia Tayassuidae está representada en América del Sur por tres
géneros: Platygonus, Catagonus y Tayassu.
La mayor cantidad de restos fósiles provienen de Argentina, Uruguay
y Brasil. Los tayasuidos (Catagonus y Tayassu) aparecen por primera
vez en Uruguay, en sedimentos posiblemente correspondientes al
Pleistoceno medio al Holoceno temprano. Platygonus se
registra en el Plioceno medio al Pleistoceno medio de Argentina,
Bolivia y Colombia.
Succinea
aff.
meridionalis.
d’Orbigny, 1837.
|
 |
|
 |
|
Ejemplar de Succenea sp.
Colección Museo de Miramar. |
|
Aspecto de un ejemplar
viviente del genero Succenea sp. |
Invertebrado, Gasterópodo.
Es un género de pequeńos caracoles terrestres,
gasterópodos pertenecientes a la familia Succineidae, que
generalmente habitan ambientes húmedos como pantanos y zonas
anegadas. El sedimento portador de estos organismos refleja la
existencia de un cuerpo de agua dulce de escasa extensión y poca
profundidad, con variaciones hídricas, desarrollado bajo un
clima cálido y húmedo dentro de la Región Neotropical, lo que
indica condiciones ambientales locales temporarias durante el
lapso estimado, hace aproximadamente 3 millones de ańos antes
del presente. Si bien sus restos han sido hallados en distintas
regiones del país, se destacan especialmente en sedimentos de la
Edad Chapadmalalense, entre las ciudades de Mar del Plata y
Miramar, así como también en la Formación Andalhuala (Plioceno
temprano) del valle de Yocavil, en la provincia de Catamarca.
Paraalbizioxylon caccavariae.
Martínez, L., 2014.
|
Algunas de los numerosas
maderas fósiles halladas en la localidad de Termas de Rió
Hondo, atribuidas al Plioceno temprano. Colección del Museo
Municipal de Paleoantropología "Rincón de Atacama". |
Paleobotánica.
La
familia Leguminosae o Fabaceae se distribuye actualmente en todos
los continentes, con excepción de la Antártida, y comprende
aproximadamente 720 géneros y unas 18.000 especies, lo que la
convierte en la tercera familia de angiospermas en cantidad de
representantes. Estos materiales constituyen el primer registro
fósil para la provincia de Santiago del Estero y permitieron obtener
valiosa información sobre el contexto ambiental de la época a partir
de leńos fosilizados hallados en la Formación Choya (Mio-Plioceno),
siendo además el primer registro fósil de maderas de leguminosas.
Los hallazgos se localizaron a orillas del río Dulce, en las
cercanías de la ciudad de Termas de Río Hondo, fueron recuperados
por Sebastián Sabater y forman parte de la colección del Museo
Municipal Rincón de Atacama. En estas investigaciones participaron
el doctor en Geología Rafael Herbert, la doctora Alexandra
Crisafulli y la profesora Mercedes Martínez, integrantes de la
Facultad de Ciencias Exactas y Naturales y Agrimensura de la
Universidad Nacional del Nordeste y del Centro de Ecología Aplicada
del Litoral (CONICET), con sede en Corrientes. Estos restos
vegetales fósiles, registrados en distintos sectores del
departamento Río Hondo, presentan troncos bien preservados y tejidos
leńosos reconocibles, lo que permitió su estudio mediante técnicas
adecuadas. Gracias a ello fue posible inferir el tipo de árboles que
integraban estos ambientes, la fisonomía de los bosques, montes o
áreas más abiertas, así como las condiciones climáticas y la
composición florística de aquel período.
Rizolitos.
Klappa, 1980.
|
 |
 |
|
Rizolitos (molde vegetales)
del Plioceno de Miramar. Colección Museo de Ciencias
Naturales de Miramar. |
Rama mandibular de un
camélido gigante (Pleistoceno) con rizolitos sobre las
mismas. Colección Museo de Miramar. |
Paleobotánica.
El término rizolito describe a aquellas rocas cuyos
rasgos estructurales, texturales y genéticos son el resultado total
o predominante de la actividad de antiguas plantas. Estas trazas de
raíces presentan morfologías y concentraciones variables, y
constituyen estructuras de bioturbación diagnósticas de procesos de
diagénesis meteórica y exposición subaérea, típicas de ambientes
continentales. El hallazgo de restos fósiles vegetales es
relativamente poco frecuente, ya que las plantas están compuestas
principalmente por celulosa, una sustancia que se descompone con
facilidad en condiciones normales. Los moldes de raíces se forman
durante la vida del vegetal, cuando bacterias y otros organismos del
suelo precipitan minerales sobre las paredes radiculares como
resultado de su actividad metabólica. Tras la muerte de la planta y
la desintegración de sus tejidos orgánicos, estas paredes
mineralizadas se rellenan con sedimentos, preservando la forma
original de la raíz. Los rizolitos pueden encontrarse asociados a
otras estructuras radiculares intrasedimentarias, como rizotúbulos y
rizoconcreciones, lo que evidencia una intensa pedogénesis. En
diversos yacimientos fosilíferos de Miramar y Centinela del Mar se
han registrado centenares de estas evidencias de origen botánico,
correspondientes tanto al Plioceno como al Pleistoceno.
Procornitermes.
Emerson, 1949.
|
 |
|
 |
|
Detalles en
restos de un Toxodontidae con marcas realizadas por
termitas. Museo Paleontológico de San Pedro. |
|
Nido de
Termitas fósiles del Plioceno de la ciudad de Miramar. Foto
del Museo Municipal Punta Hermengo. |
Insecto, Isoptero.
Es un género de termitas perteneciente a la familia
Termitidae, un grupo de insectos sociales caracterizados por la
construcción de nidos o termiteros. Se alimentan de la celulosa
contenida en la madera y sus derivados, que degradan gracias a la
acción de protozoos simbiontes presentes en su sistema digestivo. La
mayoría de las termitas habita climas tropicales y subtropicales,
aunque algunas pocas especies se adaptan a regiones templadas. En la
actualidad, la familia Termitidae se ha retirado casi por completo
de la provincia de Buenos Aires, ocupando principalmente áreas al
norte del paralelo 32, donde predominan condiciones climáticas
subtropicales. Sólo persisten pequeńas poblaciones en las sierras de
Tandil y en la península Valdés, Chubut, interpretadas como posibles
relictos de épocas pasadas. La desaparición de las termitas en la
provincia de Buenos Aires habría ocurrido durante la Edad Lujanense,
entre los 8.500 y 128.000 ańos antes del presente. Sin embargo, a
fines del Plioceno, durante la Edad Chapadmalalense de la costa
atlántica argentina, particularmente en Miramar, se han hallado
nidos fósiles de termitas y hormigas. Los géneros identificados
incluyen Procornitermes y Acromyrmex,
actualmente asociados a ambientes abiertos, húmedos, no inundables y
de clima templado. Para el Uquiense se registra el género Termes. En
2016, el Museo Paleontológico “Fray Manuel de Torres” de San Pedro
dio a conocer restos fósiles de un toxodóntido del Pleistoceno con
marcas de forma estrellada, atribuidas a la actividad alimenticia de
termitas.
Coprinisphaera. Sauer, 1955
|
 |
|
 |
|
Nido de escarabajo con
molde de la larva. Plioceno de Miramar. Museo de Ciencias
Naturales de Miramar. |
|
Aspecto de un escarabajo
pelotero. Por MarMag.2025. |
Insecto.
Las icnoespecies conocidas de Coprinisphaera
se han vinculado principalmente con escarabajos peloteros de
hábitos necrófagos, característicos de ambientes dominados por
pastizales habitados por grandes herbívoros, los cuales proporcionan
el estiércol necesario para la construcción de las bolas de cría. En
el Plioceno y el Pleistoceno bonaerense se ha recuperado evidencia
de la presencia de estos insectos mediante el hallazgo de nidos
fósiles en sedimentos fosilíferos, los cuales son raramente
identificados por los especialistas debido a su característica forma
circular. En 2014, personal del Museo de Ciencias Naturales de Punta
Hermengo de la ciudad de Miramar recuperó un nido en sedimentos del
Plioceno tardío que contenía el molde de una larva de escarabajo.
Los coleópteros presentan una enorme diversidad morfológica y ocupan
prácticamente todos los hábitats, incluidos los de agua dulce,
aunque su presencia en ambientes marinos es mínima. Son insectos
holometábolos, ya que experimentan una metamorfosis completa con
estados bien diferenciados de larva, pupa e imago, siendo frecuente
que la larva atraviese múltiples mudas. Otros icnofósiles semejantes
han sido registrados en la zona de Las Grutas, en sedimentos del
Pleistoceno superior de la ciudad de Necochea. Estudios detallados
sobre estos icnofósiles se desarrollan en el Museo Argentino de
Ciencias Naturales Bernardino Rivadavia, donde se ha determinado que
las evidencias más antiguas de este tipo se remontan al Cretácico.
Entre las icnoespecies conocidas se incluyen Coprinisphaera
akatanka, Coprinisphaera kraglievichi y
Coprinisphaera tonni.
Otros Vertebrados del Plioceno Argentino:
Sparassocynus, Pseudohalmarhipus, Nuńezia, Dankomys, Cholomys,
Proctenomys, Eucoelophorus, Pseudoplataeomys, Pithanotomys, Abrocama,
Neoepiblema, Dabbenea, Isomyopotamus, Neoreomys,
Cardiomys, Caviodon, Ocnerotherium,
Xotodon,
Xyophorus, y
Nonotherium.
(*) Imagen de autor u origen desconocido.
Ver sobre nuestras
Políticas de uso de imágenes.
|
Bibliografía Utilizada.
Acosta Hospitaleche, C., Tambussi, C.P. y
Reguero, M. 2001. Catálogo de los tipos de aves fósiles del
Museo de La Plata. Serie Técnica y Didáctica Museo de La
Plata Nro 41: 1-28
Albino A. M. 1989. Primer registro de
Colubroidae (Reptilia: Serpentes) de Argentina (Edad
Montehermorense - Plioceno) Ameghiniana. 25 (3): 281 - 287,
Buenos Aires, Argentina.
Albino, A.M. & S. Brizuela.
2009. Los reptiles escamosos de la provincia de Buenos
Aires: la perspectiva paleontológica. Boletín del Centro 6,
Actas de las Quintas Jornadas Paleontológicas Regionales:
1-21.
Ameghino, F. 1888a. Rápidas diagnosis de
algunos mamíferos fósiles nuevos de la República Argentina,
Buenos Aires, 17 pp.
Ameghino, F. 1889. Contribución al
conocimiento de los mamíferos fósiles de la República
Argentina. Actas de la Academia Nacional de Ciencias de
Córdoba, 6: 32-1028.
Benton, Michael. (1992). Dinosaurios y otros
animales prehistóricos. Ediciones Larousse Argentina S.AI.C.
Bond, M.; Cerdeńo, E.P.; López, G. 1995. Los
Ungulados Nativos de América del Sur. In Evolución climática
y biológica de la región pampeana durante los últimos cinco
millones de ańos. Un ensayo de correlación con el
Mediterráneo occidental (Alberdi, M.T.; Leone, G.; Tonni, E
P.; editores). Museo de Ciencias Naturales, Consejo Superior
de Investigaciones Científicas, Monografías, p. 259-275.
Madrid.
Burmeister, G. (1885). Examen crítico de los
mamíferos y reptiles fósiles denominados por Don Augusto
Bravard. Anales Museo Público Buenos Aires, 3 (14): 95-173.
Carlini, A. A., and G. J. Scillato-Yané.
1996. Un Euphractini (Mammalia, Dasypodidae)
del Plioceno de Chapadmalal (Buenos Aires, Argentina).
Consideraciones filogenéticas sobre los Euphractini. Rev.
del Museo de La Plata 9: 225-238.
Contreras, J.R. y Reig, O., 1965. Datos sobre
la distribución de genero Ctenomys (Rodentia, Octodontidae)
en la zona costera de la provincia de Buenos Aires
comprendida entre Necochea y Bahía Blanca. Phycis 25 (69):
169-186. Buenos Aires.
Cione, A.L.; Tonni, E.P. 1995a.
Bioestratigrafía y cronología del Cenozoico de la región
pampeana. In Evolución biológica y climática de la región
pampeana durante los últimos cinco millones de ańos. Un
ensayo de correlación con el Mediterráneo occidental. Museo
Nacional de Ciencias Naturales, Consejo Superior de
Investigaciones Científicas, Monografías, p. 47-74.
Madrid.
Creighton, G. K. 1984. Systematic studies on
opossums (Didelphidae) and rodents (Cricetidae). Ph.D.
dissertation, University of Michigan, Ann Arbor, xi+220 pp.
De Los
Reyes, M.; poiré, D; soibelzon, L; zurita, A. E; arrouy, M.
2013. First evidence of scavenging of a
Glyptodont (Mammalia, Glyptodontidae) from the Pliocene of
the Pampean region (Argentina): taphonomic and
paleoecological remarks. Paleontologia Electronica.
Ed. Coquina Press. Article number: 16.2.15A
Donadío, O.E. 1982. Restos de
anfisbénidos fósiles de Argentina (Squamata, Amphisbaenidae)
del Plioceno y Pleistoceno de la provincia de Buenos Aires.
Circ. Inf. Asoc. Paleont. Arg. 10: 10.
Esteban, G.1996. Revisión de los Mylodontinae
cuaternarios (Edentata, Tardigrada) de Argentina, Bolivia y
Uruguay. Sistemática, Filogenia, Paleobiología y
Paleozoogeografía y Paleoecología. Tesis Doctoral, Instituto
Miguel Lillo, Facultad de Ciencias Naturales, 235 p.
Tucumán.
Esteban, G.2006.
Nueva especie de
Paraeuphractus Scillato Yané (Dasypodidae-Euphractinae) del
Neógeno tardío, Noroeste de Argentina. Ameghiniana.
Francis, J.C.1965. Los géneros de la
subfamilia Mesotheriinae (Typotheria, Notoungulata) de la
República Argentina. Boletín del Laboratorio de
Paleontología de Vertebrados, Vol. 1, p. 7-31.
Gasparini, Z. y Báez, A.M. 1975. Aportes al
conocimiento de la herpetofauna terciaria de la Argentina.
Actas I Congreso Argentino Paleontología y Bioestratrigrafía,
Tucumán, 1974 2:377-415.
Gasparini Z., M. de la Fuente y O. Donadio.
(1986). Los reptiles cenozoicos de la Argentina:
Implicancias paleoambientales y evolución biogeográfica. Pp:
119-130. En: Evolución de los Vertebrados Cenozoicos de
América del Sur. Actas IV Congreso Argentino de
Paleontología y Bioestratigrafía, 2
Goin, F.J. 1991. Los Didelphoidea (Mammalia,
Marsupialia) del Cenozoico Tardío de la región pampeana.
Facultad de Ciencias Naturales y Museo de la Universidad
Nacional de La Plata, Tesis Doctoral, 323 p.
Kraglievich, J. y A Olazabal, 1959, Los
procionidos extinguidos del genero Chapadmalania Amegh. Rev.
Mus. Arg. Cienc. Nat. (Cien Zool).
Kraglievich, L. y C. Ameghino, 1940. Los
úrsidos extinguidos de Sud América. En: L. Kraglievich.
Obras de paleontología y geología, Talleres de Impresiones
Oficiales, 3: 557-627 La Plata.
M. T. Alberdi, G. Leona y E.
P. Tonni (editores). Evolución biológica y climática de la
región pampeana durante los últimos cinco millones de ańos.
Un ensayo de correlación con el Mediterráneo occidental.
Madrid, Museo Nacional de Ciencias de Madrid, Consejo
Superior de Investigaciones Científicas, 1995, Monografías
12.
M.
Magnussen Saffer, D. Boh, S. Sabater y C. Estarli.
(2015). Nuevo
registro de un proterotheriidae (Mammalia, Litopterna)
de la formación Las Cańas (Mioceno tardío - Plioceno
temprano), Termas de Rió Hondo, Santiago del Estero,
Argentina.
XXIX Jornadas Argentinas de Paleontología
de Vertebrados. Diamante, Entre
Ríos. Mayo de 2015.
Libro de Resúmenes.
Magnussen Saffer, Mariano (2015). Los Mamíferos Xenartros más
característicos del Plioceno de la Republica Argentina. Paleo
Revista Argentina de Paleontología. Boletín Paleontológico. Ańo
XIII. 129: 12-13.
Magnussen Saffer, Mariano. 2015. Aves y Reptiles representativos
durante el Plioceno de la Republica Argentina. Paleo Revista
Argentina de Paleontología. Boletín Paleontológico. Ańo XIII.
130: 11-16.
Magnussen Saffer, Mariano. 2015. Los Notoungulados más conocidos
del Plioceno de la Republica Argentina. Paleo Revista
Argentina de Paleontología. Boletín Paleontológico. Ańo XIII.
135: 25-31.
Magnussen Saffer, Mariano. 2016. Mamíferos Marsupiales y
Carnívoros representativos del Plioceno de Argentina. Paleo
Revista Argentina de Paleontología. Boletín Paleontológico. Ańo
XIV. 137: 20-25.
Marcos Cenizo,
Esteban Soibelzon & Mariano Magnussen Saffer
(2015):Mammalian predator–prey relationships and
reoccupation of burrows in the Pliocene of the Pampean
Region (Argentina): new ichnological and taphonomic
evidence, Historical Biology,DOI:
10.1080/08912963.2015.1089868.
Martínez, R. Mercedes, & Crisafulli, Alexandra. (2019).
Maderas cenozoicas de Termas de Río Hondo, Santiago del
Estero, Argentina. Revista del Museo Argentino de
Ciencias Naturales, 21(1), 87-100.
Massoia, E. y A. Fornes. 1967. El estado
sistemático, distribución geográfica y datos etoecológicos
de algunos mamíferos neotropicales (Marsupialia y Rodentia)
con la descripción de Cabreramys, género nuevo (Cricetidae).
Acta Zool. Li-lloana 23:407-430.
Marshall, L.G. 1978. Lutreolina crassicaudata.
Mammalian Species, 91:1-4.
Novas Fernando (2006). Buenos Aires hace un
millón de ańos. Editorial Siglo XXI, Ciencia que Ladra.
Serie Mayor.
Prado J.L. y Cerdeńo E. Los mamiferos
pliocenos de la fauna local Quequen Grande (provincia de
Buenos Aires, Argentina). Estudios Geol., 54: 75-83 (1998)
Pardińas, U.F.J. 1995. Capítulo 11. Los
roedores cricétidos. En: M.T. Alberdi, G. Leone y E.P. Tonni
(eds.) Evolución climática y biológica de la región pampeana
durante los últimos cinco millones de ańos. Un ensayo de
correlación con el Mediterráneo occidental , Monografías del
Museo Nacional de Ciencias Naturales, Madrid, 14: 229-256.
Pascual, R. y O. E. Odreman Rivas. 1971.
Evolución de las comunidades de los vertebrados del
Terciario argentino. Los aspectos paleozoogeográficos y
paleoclimáticos relacionados. Ameghiniana 8: 372-412.
Pascual, R.; Ortega, E.J.; Gondar, D.G.;
Tonni, E.P. 1966. Vertebrata. In Paleontografía Bonaerense.
Vertebrata I (Borrello, A.V.; editor). Comisión de
Investigaciones Científicas de la Provincia de Buenos Aires,
p. 202. Buenos Aires.
Patterson, B. y Kraglievich, L., 1960.
Sistemática y nomenclatura de las aves
fororracoideas del Plioceno argentino. Publicación del Museo
Municipal de Ciencias Naturales y Tradicional de Mar del
Plata, 1 (1): 1-52.
Quintana, Calos Adrián.
(2015). Los Fósiles de Mar del Plata. Un viaje al
pasado de nuestra región.
Fundación Félix de Azara, Vázquez Mazzini
Editores
Reguero, M.A. 1994b. Filogenia y
clasificación de los Typotheria Zittel 1893 y Hegetotheria
Simpson (Mammalia, Notoungulata). In Jornadas Argentinas de
Mastozoología (SAREM), No. 9, Resúmenes, p. 92. Vaquerías,
Córdoba.
Reguero, M.A. 1999. El problema de las
relaciones sistemáticas y filogenéticas de los Typotheria y
Hegetotheria (Mammalia, Notoungulata): análisis de los
taxones de Patagonia de la Edad-mamífero Deseadense
(Oligoceno). Universidad de Buenos Aires, Departamento de
Ciencias Biológicas, p. 1-301.
Reig, Osvaldo. (1980). Teoría
del origen y desarrollo de la fauna de mamíferos de América
del Sur. Mar del Plata, Museo Municipal de Ciencias
Naturales "Lorenzo Scaglia", 1981, Monografía Naturae, Nş 1
Reig, O. A. 1958. Notas para una
actualización del conocimiento de la fauna de la Formación
Chapadmalal. I. Lista
faunística preliminar. Acta Geol. Lilloana
2:241-253.
Reig,
O.A. Nuevos datos descriptivos sobre Chapalmatherium novum
Amegh. Revista del Museo Municipal de Ciencias Naturales y
Tradicional de Mar del Plata, 1:105-117.
Reig, O. 1958. Notas para una
actualización del conocimiento de la fauna de la Formación
Chapadmalal. II Amphiabia, Reptilia, Aves, Mammalia (Marsupialia,
Didelphidae, Boriiyaenidae). Acta Geol. Lilloana 2: 255-283.
Rusconi, C., 1958. Ave fosil del Plioceno de
Buenos Aires. Revista Museo Historia Natural, 21: 1-3.
S
Brizuela, AM Albino. Los reptiles escamosos del Plioceno de
la costa atlántica entre Mar del Plata y Miramar, provincia
de Buenos Aires, Argentina. Revista del Museo Argentino de
Ciencias Naturales 14 (1), 47-56
Scillato-Yané, G.J.; Carlini, A.A.; Vizcaíno,
S.F.; Ortíz Jaureguizar, E. 1995. Los Xenarthros. In
Evolución biológica y climática de la región pampeana
durante los últimos cinco millones de ańos. Un ensayo de
correlación con el Mediterráneo occidental (Alberdi, M.T.;
Leone, G.; Tonni, E.P.; editores). Museo de Ciencias
Naturales, Consejo de Investigaciones, Monografías, p.
183-209. Madrid.
Straccia, P.C. (1999).
Anatomía funcional de Argyrolagus scagliai (Mammalia:
marsupialia: argyrolagidae) del Plioceno en la Argentina.
Tesis doctoral.
Facultad de Ciencias
Naturales y Museo.
Tambussi, C. 2001. Hermosiornis
milneedwarsi. Aire libre 2 (9):106.
Tambussi, C. P., 1995. Aves. En: Alberdi, M.
T., G. Leone y E. P. Tonni (eds.), Evolución biológica y
climática de la Región Pampeana durante los últimos 5
millones de ańos. Monografía del Museo Nacional de Ciencias
Naturales, CSIC, Espańa, cap. 7: 145-161.
Tonni. E. P., 1980. The present state of
knowledge of the Cenozoic birds of Argentina. In: Papers in
avian paleontology honoring Hildegard Howard, K. Cambell,
ed., Contributions in Science.
Natural History Museum of Los Angeles
Country, 330: 105-114.
Zárate, M. A. 1989.
Estratigrafía y geología del Cenozoico tardío aflorante en
los acantilados marinos comprendidos entre Playa San Carlos
y el arroyo Chapadmalal, Partido de General Pueyrredón,
Provincia de Buenos Aires. Tesis doctoral inédita. Facultad
de Ciencias Naturales y Museo, Universidad Nacional de La
Plata, 221 p.
Ver mas
bibliografía utilizada para hacer la presente Pagina;
AQUÍ.
Ver bibliografía de divulgación sugerida;
AQUÍ. |
|