|
|
|
Fósiles del Pleistoceno
de Argentina: |
|
|
El
Periodo/Época Pleistoceno abarca los desde los 2,5 millones de ańos
hasta los 11,7 mil ańos antes del presente. Se halla representado por
cuatro edades.

Sedimentos del Pleistoceno de Miramar.
|
 |
La edad
Marplatense
(anteriormente denominada
Uquiense)
se ubica en un lapso transicional dentro del Plioceno y
abarca aproximadamente entre los
3,3 y 1,9 millones
de ańos. Sus afloramientos pueden observarse en las
barrancas costeras comprendidas entre las ciudades de
Mar del Plata y
Miramar. La edad
Ensenadense
corresponde a sedimentos con una antigüedad aproximada de
1,8 millones a 700
mil ańos, mientras que la edad
Bonaerense
abarca un intervalo comprendido entre
700 mil y 130 mil
ańos antes del presente. La última edad del
Pleistoceno es la
Lujanense, con una cronología que se extiende desde
130 mil hasta 8 mil
ańos, marcando la transición hacia el
Holoceno,
período en el cual se produce la extinción de gran parte de
los grandes mamíferos y sobreviven las especies actuales.
Todas estas edades se encuentran representadas en las
formaciones geológicas comprendidas entre
Santa Clara del
Mar y Monte Hermoso, en la provincia de
Buenos Aires,
Argentina, cuyo registro fosilífero y
estratigráfico constituye uno de los más importantes del
Cenozoico a nivel
mundial. Durante los últimos
dos millones de
ańos, el nivel del mar experimentó un marcado
descenso hasta alcanzar su posición actual, como resultado
de diversos eventos significativos. En este período
habitaron algunas de las criaturas más grandes y singulares
de toda la Era Cenozoica. Sin embargo, este intervalo
presenta una paradoja notable: la extinción de
aproximadamente el
96 % de la megafauna sudamericana, junto con
especies de origen norteamericano, ocurrida en un breve
lapso entre 12 y
10 mil ańos atrás. |
|
La causa de este proceso de
extinción aún no se conoce con certeza, aunque una de las
hipótesis más debatidas es la llegada de una especie
invasora y de locomoción bípeda:
Homo sapiens.
Estos grupos humanos pudieron haber cazado a los grandes
mamíferos, aunque no existen evidencias concluyentes de
cacerías masivas, selectivas o sistemáticas que permitan
seńalar al ser humano como la principal amenaza para estas
especies. Los
cambios climáticos abruptos también habrían
desempeńado un papel relevante, ya que existen numerosos
registros de un evento climático agudo ocurrido entre
8 y 6 mil ańos atrás.
Prácticamente todas las regiones del país conservan
sedimentos correspondientes al
Pleistoceno,
aunque algunas localidades presentan una riqueza fósil
significativamente mayor que otras.
<<<
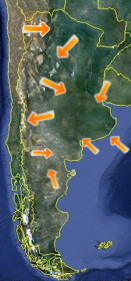
Principales sitios
fosilíferos de Argentina. |
Salvator (Tupinambis) teguixin.
Daudin, 1802.
|
 |
|
 |
|
Esqueleto
recreado in situ Tupinambis. |
|
Reconstrucción en vivo del genero Tupinambis sp
del Pleistoceno pampeano.
Por MarMag.2025. |
Reptil
Squamata.
Es el lagarto de mayor tamańo de la familia
Teiidae, con un
registro que se remonta al
Mioceno temprano de la Patagonia y una presencia continua en
Argentina a lo largo de todo el Mioceno, extendiéndose hasta el
Holoceno–Reciente. Los
reptiles escamosos (Squamata)
constituyen el segundo grupo más numeroso de vertebrados terrestres, con
más de 8.200 especies,
superados únicamente por las aves. No obstante, pese a su notable
diversidad y abundancia actual, han sido históricamente relegados en la
literatura paleontológica por diversas razones. En la actualidad,
Tupinambis está
representado por al menos seis
especies: T. duseni,
T. longilineus,
T. merianae,
T. quadrilineatus,
T. rufescens y
T. teguixin, todas restringidas
al territorio sudamericano, al este de la cordillera de los Andes.
Recientemente, se describió una forma extinta,
Tupinambis uruguaianensis,
procedente del Pleistoceno de
Brasil, de mayor tamańo que cualquiera de las especies
vivientes. En 1914, Rovereto
describió cuatro especies fósiles del género (T.
preteguixin, T. prerufescens,
T. brevirostris y
T. multidentatus),
provenientes de Monte Hermoso,
a unos 60 km al este de Bahía Blanca, sobre la costa atlántica del sur
de la provincia de Buenos Aires. Su registro paleontológico en
sedimentos pampeanos se extiende desde el
Mioceno tardío hasta el
Holoceno temprano, aunque su mayor abundancia se registra hacia
el final del Plioceno,
bajo condiciones climáticas predominantemente cálidas. Este lagarto se
caracteriza por la marcada diferenciación de su dentición, que incluye
incisivos, dientes con forma de caninos y otros aplanados, similares a
molares. Podía superar el
metro de longitud total, aunque la cola representaba más de la
mitad del cuerpo. Los machos presentaban una cabeza más ancha, producto
del mayor desarrollo de la musculatura mandibular. Su dieta incluía
anfibios, gusanos y otros reptiles, así como pequeńos mamíferos, entre
ellos cricétidos y algunos marsupiales. Ocasionalmente, sus restos
aparecen articulados dentro de antiguas
paleocuevas,
excavadas por el propio Tupinambis
o por otros vertebrados y posteriormente ocupadas por este reptil.
Importantes hallazgos para la paleofauna pampeana proceden del
Plioceno del arroyo Las
Brusquitas y la Barranca de Los Lobos, así como de sedimentos
más recientes, incluidos
sitios arqueológicos. En la región pampeana, se lo registra en
las formaciones Monte Hermoso,
Chapadmalal y Vorohué. Estudios recientes proponen reemplazar
la asignación al género Tupinambis
por Salvator sp.,
dado que los ejemplares fósiles resultan
indistinguibles de las
especies actuales.
Epicrates
sp. Wagler, 1830.
|
 |
|
 |
|
Vértebras
halladas en el Pleistoceno de Termas de Rió Hondo. Museo
Municipal Rincón de Atacama. |
|
Aspecto de la
boa del genero Epicrates.
Por MarMag.2025. |
Reptil, Ofideo. Es
un género de serpientes perteneciente a la familia de las boas,
representado en la actualidad por
Epicrates alvarezi, una especie que habita regiones cálidas y
semiáridas del centro de América del Sur. El estudio de estas serpientes
gigantes, emparentadas con los antepasados de anacondas, boas y pitones,
permite comprender no solo la historia evolutiva del grupo en
Sudamérica, sino también los eventos paleobiogeográficos que modificaron
de manera significativa la fisonomía del continente y su fauna.
Epicrates es
endémica de las ecorregiones terrestres del Chaco occidental y del Chaco
árido, cuya vegetación original —hoy severamente reducida por la
expansión agropecuaria— corresponde a los distritos fitogeográficos
occidental y árido de la provincia fitogeográfica chaqueńa. Se considera
que su representante fósil, que habitó esta región hace más de 20 mil
ańos, ocupaba ambientes muy similares a los actuales. En la actualidad,
la boa arco iris se distribuye principalmente en las provincias de
Catamarca, Chaco, Córdoba, Formosa, Jujuy, La Rioja, Salta, San Luis,
Santa Fe, Santiago del Estero y Tucumán. Como todas las serpientes, era
estrictamente carnívora y presentaba dientes agudos y recurvados en la
mandíbula superior, el paladar y la mandíbula inferior; capturaba y
mataba a sus presas por constricción. De hábitos nocturnos y
comportamiento tímido, se alimentaba principalmente de pequeńos
mamíferos, complementando su dieta con aves. Se desplazaba mayormente
por el suelo, aunque podía trepar a los árboles ante situaciones de
peligro o para refugiarse en cavidades de troncos antiguos. En el
registro fósil, el género fue identificado por primera vez en sedimentos
del Pleistoceno, a unos 10 kilómetros de la ciudad de Termas de Río
Hondo, a partir de restos de la columna vertebral actualmente
resguardados en el Museo Municipal Rincón de Atacama, los cuales fueron
estudiados por Pablo Gaudioso y Adriana Albino.
Rhinella arenarum.
Hensel, 1867.
|
 |
|
 |
|
Esqueleto parcialmente completo y recreado de
Rhinella. |
|
Aspecto de
Rhinella sp durante el Pleistoceno. (*). |
Anuro.
Los bufónidos se caracterizan por carecer de dientes y poseer glándulas
parotoideas ubicadas en la parte posterior de la cabeza, las cuales
secretan toxinas con distintos efectos defensivos. Como todos los
anuros, atraviesan un proceso de metamorfosis, iniciando su ciclo vital
como renacuajos de aspecto pisciforme, sin extremidades y con
respiración branquial, para luego desarrollar patas, reabsorber la cola
y adoptar la respiración pulmonar. Dentro de esta familia cosmopolita de
anuros neobatracios se incluye el género
Rhinella, nativo del
sur de Norteamérica, América Central y América del Sur, cuyas especies
fueron originalmente agrupadas dentro del género
Bufo, posteriormente
dividido en Rhinella y
Rhamphophryne. El
registro fósil del género
Rhinella se remonta al
Oligoceno de Bolivia y al
Mioceno medio de La Venta,
Colombia, aunque aún se requieren mayores evidencias para
reconstruir con precisión la historia evolutiva de la familia durante el
Terciario. Su hábitat comprendía cavidades próximas a cuerpos de agua y
áreas con vegetación abundante, aunque su notable capacidad de
adaptación le permitió ocupar también ambientes semiáridos. Presentaban
hábitos alimentarios oportunistas, consumiendo pequeńos vertebrados e
incluso individuos juveniles de su misma familia. En la provincia de
Buenos Aires, se han hallado restos asignables a este género en
sedimentos del Plioceno
tardío, Pleistoceno y Holoceno, si bien la fragilidad de sus
huesos ha limitado su preservación y, en consecuencia, la abundancia del
registro fósil disponible.
Odontophrynus
sp. Reinhardt & Lütken, 1862.
| |
|
 |
|
|
|
Aspecto del pequeńo
escuerzo del Pleistoceno, Odontophrynus sp.
|
Anuro. Es
un género de anfibios anuros perteneciente a la familia
Odontophrynidae, cuyos representantes son conocidos comúnmente
como escuercitos; su registro fósil es escaso y se conoce principalmente
a partir del Ensenadense,
lo que sugiere que no presentaba diferencias significativas respecto de
las especies actuales. Probablemente excavaba con las extremidades
posteriores y permanecía oculto la mayor parte del tiempo, emergiendo de
sus refugios tras lluvias intensas para alimentarse con mayor actividad
y reproducirse. Alcanzaba un tamańo aproximado de
7 centímetros, con un
cuerpo robusto y globoso, extremidades cortas y una cabeza ancha y
redondeada. Habitaba lagunas permanentes de aguas relativamente
profundas utilizadas para la reproducción, muchas veces cubiertas por
una densa vegetación flotante. En 2024, un equipo integrado por
investigadores del Museo de Ciencias Naturales de Miramar, la Fundación
Azara y el Laboratorio de Anatomía Comparada y Evolución de los
Vertebrados (MACN–CONICET) recuperó egagrópilas con restos de
microvertebrados, entre los cuales se identificaron pequeńos fósiles
asignables a Odontophrynus sp.,
correspondientes al menos a tres individuos hallados en sedimentos
pleistocenos de la Reserva Natural Provincial de
Centinela del Mar;
actualmente, en esta misma región habita el
Escuercito común (Odontophrynus
americanus).
Chelonoidis lutzae.
Zacarías et al., 2013.
 |
 |
 |
|
Pieza tipo e Chelonoidis
letzae,
en el Museo Paleontológico de Topori, Corrientes.
|
Placa de
Chelonoidis
del Plioceno de Entre Ríos. Museo de La Plata. |
Aspecto de Chelonoidis. Escultura de Gabriela
Alejandra Monteverde, realizada para la Fundación Azara. |
|
 |
 |
 |
| Enormes
caparazones de Chelonoidis
letzae,
en el Museo Paleontológico de Topori, Corrientes.
|
Geochelone carbonaria,
del Pleistoceno del
arroyo Barrenechea,
Diamante, Entre Ríos. Hallada por
César Travieso .
Foto de Raul Kemer. |
Caparazón
comparable con las tortugas gigantes de Galápagos. Este
fósil de Chelonoidis sp.
es el Pleistoceno del Arroyo El
Cura. Museo Almeida, Gualegauychu. |
Reptil
Testudinido. Las
tortugas constituyen un grupo singular de reptiles, caracterizados
por poseer la cabeza, las extremidades y la cola protegidas por un
caparazón dorsal y un plastrón ventral, y su estudio paleontológico
ha permitido reconstruir la anatomía y el modo de vida de las
tortugas terrestres gigantes que habitaron la provincia de
Corrientes durante el Pleistoceno. Investigaciones realizadas por
equipos de la UNNE, a partir del análisis de registros fósiles,
aportaron información clave sobre los ambientes de aquel período y
los cambios naturales que condujeron a la desaparición de gran parte
de la megafauna. Las tortugas terrestres presentan caparazones más
globosos y una reducción en el número de falanges de los dedos
—generalmente de tres a dos—, una adaptación que les permite
soportar mejor el peso corporal. En 2008 se rescataron ejemplares
fósiles de una tortuga gigante en la localidad de Bella Vista,
procedentes de sedimentitas de la Formación Toropí, con una
antigüedad estimada entre 50.000 y 35.000 ańos (Pleistoceno tardío),
sumándose a hallazgos previos como el primer ejemplar recuperado por
Beatriz Álvarez y Rafael Herbst, y un segundo, más completo aunque
fragmentario, extraído por el grupo de Paleontología del Centro de
Ecología Aplicada del Litoral, ambos asignados al género
Chelonoidis.
En 2009 se confirmó definitivamente la presencia de este género a
partir de un ejemplar más completo exhumado en 2007 por
investigadores de la FACENA–CECOAL y de la Universidad de La Plata.
El análisis ecomorfológico de un húmero grácil de una tortuga
gigante terrestre de Bella Vista permitió inferir que estas especies
habitaban áreas abiertas bajo condiciones climáticas más frías y
áridas que las actuales. A diferencia de las tortugas gigantes de
las islas Galápagos, cuyo gran tamańo se explica por la ausencia de
depredadores, las formas continentales fósiles desarrollaron
caparazones menos abiertos y notablemente más gruesos como defensa
frente a grandes carnívoros, entre ellos lobos de gran porte y
felinos como los tigres dientes de sable. Recientemente, Diego
Gambetta, del Museo de Mar de Ajó, y Federico Agnolín, del Museo
Argentino de Ciencias Naturales y la Fundación Azara, documentaron
la presencia del género
Chelonoidis
en la localidad de Mar de Ajó, provincia de Buenos Aires, a partir
de una placa marginal, correspondiente a un período en el que la
región presentaba un ambiente y clima similares a los patagónicos.
Asimismo, en 2022, integrantes del Laboratorio de Anatomía Comparada
y Evolución de los Vertebrados (LACEV), durante la revisión de
materiales fósiles del Museo Provincial Carlos Ameghino de la ciudad
de Mercedes, estudiaron restos de caparazón hallados en el río
Luján, en niveles de aproximadamente 100.000 ańos de antigüedad, y
describieron una nueva especie,
Chelonoidis petrocellii,
del Pleistoceno de la provincia de Buenos Aires, de tamańo
comparable a las especies actuales pero con un caparazón
considerablemente más grueso.
Pampagyps imperator.
Agnolin y Brissón Egli, 2017.
|
 |
|
 |
|
Metatarso atribuido a
Pampagyps imperator. Prensa. |
|
Aspecto de Pampagyps.
Ilustración Gabriel Lio. Prensa. |
Ave,
Cathartidae.
El hallazgo es excepcional no solo para la provincia de
Buenos Aires, sino para toda la Argentina, ya que constituye el
primer registro de un cóndor extinto hasta ahora desconocido y
preservado en un estado notable. Este imponente ave alcanzaba más de
2,5 metros de envergadura
alar, poseía garras considerablemente más robustas que las
del cóndor actual y, según las estimaciones de los paleontólogos,
habría sido capaz de capturar presas vivas. Sus restos fósiles
fueron descubiertos en el partido de
Marcos Paz, a tan
solo 34 kilómetros de la Ciudad de Buenos Aires, y su nombre puede
interpretarse como “buitre emperador de las Pampas”. El análisis
anatómico reveló una gran capacidad de apertura y cierre de las
garras, evidenciada en un hueso del miembro posterior con
inserciones musculares mucho más desarrolladas que en los cóndores
modernos, lo que refuerza la hipótesis de un comportamiento
predador. A diferencia de los cóndores actuales, que se alimentan
casi exclusivamente de carrońa, este cóndor emperador pampeano, con
una envergadura que pudo alcanzar los
3 metros, habría
representado una amenaza para pequeńos vertebrados, como roedores,
reptiles y aves acuáticas, entre ellas patos.
Teratornis sp.
Miller, 1909.
|
 |
|
 |
|
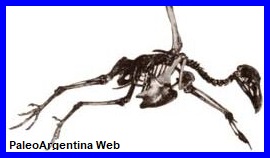
Aspecto del esqueleto de Teratornis. |
|
Aspecto de Teratornos.
Por MarMag.2025. |
Ave,
Teratorno.
Los teratornos fueron aves voladoras
gigantes de hábitos carrońeros y predadores que habitaron el
continente americano durante casi
25 millones de ańos,
hasta su extinción ocurrida hacia el final del
Pleistoceno, hace unos 12
mil ańos. En Argentina, sus restos han sido hallados en
Santa Fe, Pehuen Co y en las inmediaciones de Miramar; si bien no se
ha podido determinar con precisión el género, los investigadores
consideran que podrían corresponder a
Teratornis.
Estas aves superaban en tamańo a los cóndores actuales y hoy se
encuentran completamente extintas; se reconocen alrededor de
siete especies de
teratornos, entre ellas
Argentavis magnificens, el ave voladora más grande
conocida hasta el momento, con un peso estimado de
70 kilogramos y una
envergadura alar de hasta 7
metros, que vivió en el Mioceno de Argentina. Los primeros
restos de teratornos fueron descubiertos en 1909 en los célebres
pozos asfálticos de Rancho
La Brea, en California, donde se recuperaron centenares de
ejemplares de la especie
Teratornis merriami,
que dio nombre a esta familia de aves extintas. Se considera que
los teratornítidos se originaron en
América del Sur,
ya que sus registros más antiguos provienen de yacimientos de Brasil
y Argentina con edades comprendidas entre
25 y 5 millones de ańos;
tras este intervalo desaparecen del registro fósil sudamericano,
pero se vuelven notablemente abundantes y diversos en
América del Norte
hasta su extinción definitiva. La ausencia de estas aves gigantes en
América del Sur durante los últimos cinco millones de ańos ha
constituido un enigma, y la escasa evidencia fósil disponible impide
por el momento diferenciar con certeza los restos sudamericanos de
las especies norteamericanas, aunque
Teratornis
habría alcanzado una envergadura cercana a
4 metros, una
altura aproximada de 80
centímetros y un peso de alrededor de
15 kilogramos.
Rhea fossilis.
Ameghino, 1882.
|
 |
 |
 |
|
Cáscaras de
huevo del genero
Rhea del ensenadense. Museo Punta Hermengo. |
Huellas
fósiles de
Aramayoichnus
en el Pleistoceno de Miramar. |
Aspecto de
Rhea fossilis.
Por MarMag.2025. |
Ave.
Rheiformes.
Los Rheidae
constituyen una familia endémica de la región neotropical, de origen
sudamericano, que aparentemente nunca abandonó el continente y cuyo
registro se extiende desde el
Mioceno hasta el Holoceno.
Se trata de aves de gran tamańo, exclusivamente terrestres y
corredoras, con capacidad de vuelo nula, cuya presencia en el
registro fósil es indicativa de
ambientes abiertos,
dominados por pastizales y con escasa cobertura boscosa. Aunque sus
restos óseos son poco frecuentes, se han identificado
huellas fósiles
atribuidas a sus pisadas tanto en
Pehuen Co como en
sedimentos del Pleistoceno y
Holoceno de Miramar, asignadas al icnogénero
Aramayoichnus.
En esta última localidad, personal del Museo de Ciencias Naturales
de Miramar recuperó además un fragmento de sedimento con
cáscaras de huevo
atribuibles a esta familia, correspondientes al
Ensenadense, con
una antigüedad aproximada de
700 mil ańos.
Rhea fossilis representa
el único réido extinto conocido del
Pleistoceno tardío,
y habría convivido con otras formas aún vivientes o registradas en
el pasado reciente, como Rhea
subpampeana, Rhea
pampeana, Rhea
anchorenensis, Rhea
americana y Pterocnemia
pennata, aunque la
sinonimia dentro del género
Rhea continúa siendo objeto de debate.
|
Toxodon platensis. Owen,
1837. |
|
toxodon platensis toxodon platensis toxodon platensis
toxodon platensis toxodon platensis toxodon platensis
toxodon platensis |
|

|
|

|
|
Esqueleto de
Toxodon platensis en el Museo de Ciencias Naturales de
Miramar, montado por la Fundación Azara. |
|
Reconstrucción
de Toxodon platensis. (*). |
|
 |
 |
|
Esqueleto de
Toxodon
en el Museo Argentino de Ciencias Naturales, Buenos Aires
(MACN). |
Cráneo y
mandíbula hallado en Mar del Sud, exhibido en el Museo de Ciencias Naturales de Miramar. |
Mamífero
Notoungulado. El nombre
Toxodon significa
“diente en arco” y su evolución estuvo estrechamente ligada al
prolongado aislamiento geográfico de América del Sur, que favoreció
el desarrollo de mamíferos singulares capaces de diversificarse y
ocupar distintos ambientes. Este robusto herbívoro alcanzaba cerca
de 3 metros de longitud
y una masa estimada de una
tonelada; su cráneo medía alrededor de
70 centímetros y
poseía mandíbulas provistas de incisivos de crecimiento continuo,
con esmalte dispuesto en bandas longitudinales. Aunque su aspecto
recuerda al de los hipopótamos y rinocerontes actuales, no existe
parentesco directo, tratándose de un claro ejemplo de
evolución convergente,
en la que linajes no emparentados desarrollan rasgos similares al
ocupar nichos ecológicos semejantes. Tradicionalmente se lo ha
asociado a llanuras
abiertas y ambientes pantanosos, donde se alimentaba
principalmente de vegetación que podía arrancar mediante sus
mandíbulas en forma de “pala”; sin embargo, otros autores proponen
que también habitó
ambientes semiáridos, ya que sus restos han sido
registrados en sedimentos eólicos del
Pleistoceno tardío,
como en Sayape (provincia de San Luis) y Caleufú (La Pampa).
Asimismo, en la localidad de
Tres Arroyos,
provincia de Buenos Aires, se documentó la presencia del género
Toxodon
asociada a restos culturales en un sitio arqueológico con una
antigüedad cercana a 8.500
ańos antes del presente. En 1916 se dio a conocer el
hallazgo de un fémur de
Toxodon con una punta de proyectil incrustada, descubierto en
la ciudad de Miramar,
hecho que generó intensas controversias científicas y que hoy es
considerado un fraude de la época. El género presenta una amplia
distribución estratigráfica, desde el
Plioceno tardío hasta el
Holoceno temprano, y una extensa distribución geográfica
que abarca desde Venezuela
hasta el sur de la provincia de Buenos Aires, con registros
también en Paraguay,
Brasil y Bolivia. Entre las especies reconocidas se
incluyen Toxodon
chapalmalensis, T.
darwini, T. ensenadensis,
T. platensis,
T. gezi y
T. gracilis, aunque
algunos autores sostienen que
T. platensis y T. gezi
podrían corresponder en realidad a una misma especie, cuestión que
aún requiere una revisión sistemática integral del género.
|
Macrauchenia patachonica.
Owen, 1838. |
|
Macrauchenia
patachonica Macrauchenia patachonica Macrauchenia
patachonica Macrauchenia patachonica Macrauchenia |
|
 |
|
 |
|
Esqueleto de
Macrauchenia
en el MACN. (Museo Argentino de Ciencias Naturales de
Buenos Aires). |
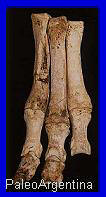
Pata de tres dedos. (*) |
Reconstrucción de
Macrauchenia patachonica (*). |
|
 |
|
 |
|
Cráneo y Mandíbula de Macrauchenia patachonica. Museo
de Ciencias Naturales de Miramar. Gentileza de Adrián
Giacchino. |
|
Huella de Eumacrauchenichnus
patachonicus, en el Sitio
Paleoicnologico de Miramar. Hallazgo Mariano Magnussen. |
Mamífero
Litopterno.
Los primeros restos de
Macrauchenia
fueron descubiertos por
Charles Darwin durante su histórico viaje a bordo del
Beagle a comienzos de la
década de 1830, en sedimentos del
Pleistoceno tardío
aflorantes en los alrededores de
Punta Arenas, en el
extremo sur de la Patagonia, y dados a conocer científicamente por
el paleontólogo inglés
Richard Owen en 1838. De aspecto singular y casi propio de
la ciencia ficción, su nombre significa “cuello largo” y pertenece
al orden Litopterna;
alcanzaba dimensiones comparables a las de los camellos actuales,
pero presentaba los orificios nasales desplazados hacia arriba y una
gran fosa elíptica que indica la presencia de una trompa,
probablemente algo más larga que la del tapir moderno. Poseía patas
largas con pies ungulados provistos de
tres dígitos,
aunque es probable que no fuera un corredor veloz, ya que las
proporciones de sus extremidades posteriores se asemejan a las de
animales no cursoriales, como la jirafa. Su dieta estaba basada en
vegetación de ambientes
pantanosos y en el pastoreo, y aunque algunos autores han
propuesto una adaptación a un modo de vida semiacuático, sus restos
se han hallado tanto en depósitos vinculados a cuerpos de agua como
en sedimentos eólicos de
ambientes áridos y semiáridos. Presentaba molares
superiores de sección cuadrangular y
coronas altas, y
se extinguió hacia fines del Pleistoceno y comienzos del Holoceno.
En la actualidad, los hallazgos fósiles de
Macrauchenia son escasos
en América Latina;
Macrauchenia patachonica se registra en sedimentos
pleistocenos de Brasil,
Paraguay, Chile, Bolivia, Uruguay y Argentina, mientras que
sus antecesores más antiguos aparecen en el
Montehermosense (Plioceno),
representados por
Promacrauchenia antiqua. Asimismo, se conocen restos
relativamente recientes procedentes de la
Cueva del Mylodon,
con una antigüedad cercana a
11.000 ańos, y
del Sitio 2 de Arroyo Seco,
en la provincia de Buenos Aires, con unos
8.000 ańos. Entre
las especies relacionadas se incluyen
Epitherium laternarium,
Eoauchenia primitiva,
Diplasiotherium robustum,
Neolicaphrium recens,
Promacrauchenia kraglievichi,
Promacrauchenia chapadmalense,
Windhausenia delacroixi,
Pseudomacrauchenia yepesi
y Macraucheniopsis
ensenadensis.
Neolicaphrium recens.
Frenguelli,
1921.
|
 |
|
 |
|
Fragmento de
mandíbula de Neolicaphrium, procedente del Pleistoceno de
Termas de Rió Hondo, Santiago del Estero |
|
Posible
aspecto del
proteroterio
Neolicaphrium recens. MarMag.2026. |
Mamífero
Litopterno.
Se trata de un mamífero de pequeńo tamańo,
superficialmente similar a los caballos actuales pero sin parentesco
alguno, perteneciente a la familia de los
proteroteríidos, un
linaje muy primitivo originado en América del Sur. Durante cerca de
50 millones de ańos de
relativo aislamiento, los mamíferos sudamericano-antárticos
evolucionaron en un verdadero continente-isla, dando lugar a formas
únicas como Neolicaphrium
recens, el último representante conocido de los
proteroteríidos. La expansión de las planicies abiertas en gran
parte del continente sudamericano favoreció la evolución de animales
corredores de estructura liviana; los primeros proteroterios
presentaban un dedo central muy desarrollado en cada pie y dos
laterales reducidos que apenas tocaban el suelo, una condición
comparable a la observada en las patas traseras de los jabalíes
actuales. Estos pequeńos “falsos caballos” eran gráciles, de dorso
relativamente corto y extremidades alargadas con pezuńas; el cráneo
mostraba un rostro no particularmente alargado y ojos grandes,
recordando más a gacelas u otros herbívoros pequeńos o medianos que
a los caballos modernos. Las proporciones de sus miembros sugieren
que habitaban ambientes
más boscosos, lo que permite realizar inferencias
paleoambientales y paleoclimáticas. Recientemente se dio a conocer
la presencia del género
Neolicaphrium
en sedimentos del
Pleistoceno de Termas de Río Hondo, en la provincia de
Santiago del Estero, donde previamente ya se había registrado parte
de un esqueleto de otro proteroterio en niveles
Mio-pliocenos;
además, se conocen escasos restos de
Neolicaphrium procedentes
del Pleistoceno de
Corrientes, Córdoba, Santa Fe y Uruguay.
Mesotherium cristatum.
Serres, 1967.
Mamífero
Notoungulado.
Fue un género de
mesoungulados notoungulados
de hábitos estrictamente terrestres y alimentación herbívora, basada
principalmente en follajes duros presentes en
llanuras abiertas.
Alcanzaba un tamańo comparable al de una oveja actual y poseía
extremidades largas y esbeltas, adaptadas para la carrera a gran
velocidad, una estrategia defensiva frente a depredadores como los
feroces marsupiales carnívoros y las gigantescas aves que habitaban
Sudamérica en esa época. Su cuerpo era delgado pero robusto, y su
cráneo se diferenciaba notablemente de otros herbívoros similares
por presentar una dentición
de tipo rodentiforme. Representa un grupo relativamente
moderno dentro de los
tipoterios, exclusivo del continente sudamericano, cuyos
restos aparecen con frecuencia desde fines del
Plioceno medio
(posiblemente
Pseudotypotherium) hasta el
Pleistoceno medio–superior,
especialmente en los barrancos del litoral marítimo bonaerense entre
Miramar y Mar del Plata,
aunque son más abundantes en localidades como
Ensenada y Olivos.
Entre los géneros relacionados se incluyen
Protypotherium antiquum,
Typotheriopsis chasicoensis,
Protypotherium australe,
Pseudotypotherium histatum
y Mesotherium cristatum.
|
Megatherium americanum.
Cuvier, 1796. |
|
megatherium americanum megatherium americanum megatherium
americanum megatherium americanum megatherium |
|
|
|
Esqueleto
montado en el Museo Argentino de Ciencias Naturales de
Buenos Aires "Bernardino Rivadavia". Al lado, uno de los dos
esqueletos de Megaterio exhibidos en el Museo de La Plata.
El técnico Mariano Magnussen Saffer junto a un cráneo de
Megaterio en plena preparación en el Museo de Ciencias
Naturales de
Miramar, hallado por Pablo Lamego. |
|
 |
 |
 |
|
Huellas de
Megatherium en Pehuen Co. Fuente; Revista
Ciencia Hoy. |
Huellas del
icnogenero
Neomegatherichnum
en Pehuen Co. |
Reconstrucción de Megaterio. |
|
Mamífero
Pilosa.
En la actualidad, los
Pilosa están
representados por mamíferos pequeńos y medianos, como osos
hormigueros, osos meleros y perezosos de tres dedos, caracterizados
por hábitos arborícolas, movimientos lentos y un metabolismo bajo;
sin embargo, durante el
Pleistoceno habitaron en Sudamérica formas gigantes, entre
las que destacó Megatherium,
descrito por primera vez en 1796 por el naturalista francés
Georges Cuvier a
partir de un esqueleto completo hallado en 1789 en las barrancas del
río Luján
(provincia de Buenos Aires). Su nombre, que significa “bestia
grande”, refleja adecuadamente que fue uno de los mayores mamíferos
terrestres que vivieron en la región pampeana y en el Pleistoceno
continental sudamericano, constituyendo, junto con
Glyptodon, uno de los
elementos más conspicuos de la megafauna de ese período. Presentaba
una dentición compuesta por molariformes en forma de columnas
prismáticas cuadrangulares y tradicionalmente se lo ha considerado
herbívoro, aunque se han propuesto dietas mixtas, carrońeras o
incluso carnívoras, a partir de evidencias como marcas pares en
huesos de megafauna hallados en Uruguay, compatibles con su
dentición, y estudios isotópicos que sugieren consumo de carne en
algunos perezosos gigantes extintos, sin poder determinar si fue por
caza o carrońa. El registro del género se extiende desde el
Vorohuense hasta
el Lujanense temprano
(Megatherium americanum)
y tardío (Megatherium sp.),
con una distribución geográfica restringida al sur de América del
Sur —Argentina, Brasil, Uruguay, Paraguay, Chile, Perú y Bolivia—,
siendo reemplazado hacia el norte por el género
Eremotherium,
de rasgos más primitivos y probablemente adaptado a climas más
cálidos. Megatherium
podía superar los 4,5
metros de altura al erguirse sobre sus patas traseras,
apoyándose en la cola a modo de trípode para alcanzar el follaje
arbóreo y obtener una amplia visión del entorno, y su peso estimado
rondaba las cinco
toneladas; su cuerpo estaba cubierto por una densa pelambre
y sus extremidades anteriores, largas y robustas, poseían grandes
garras que facilitaban el acceso a la vegetación. En
Pehuen Co
(provincia de Buenos Aires) se identificó un importante yacimiento
paleoicnológico con huellas de este animal, de unos
11.000 ańos de
antigüedad, asociadas a rastros de otras especies extintas y
actuales, y se sabe que coexistió con grupos humanos en la provincia
de Buenos Aires hace aproximadamente
8.500 ańos. En
2018 se dio a conocer el hallazgo de un fémur y una mandíbula de
Megatherium filholi
en la ciudad de Miramar,
una forma de menor tamańo descrita originalmente por
Francisco Pascasio Moreno
hace más de 130 ańos, lo que permitió revalidar la especie, y
posteriormente se reportó un ejemplar juvenil de
Megatherium sp. del
Plioceno, cercano
a Miramar, que junto con otros registros de Bolivia y Perú
constituye uno de los más antiguos conocidos para el género; entre
las formas relacionadas se incluyen
Plesiomegatherium,
Megalonychops carlesi,
Pronothrotherium typicum
y Diheterocnus sanmartini.
|
Mylodon
darwinii. Owen, 1839. |
|
mylodon darwinii mylodon darwinii mylodon darwinii mylodon
darwinii mylodon darwinii mylodon darwinii mylodon darwinii |
|
 |
 |
|
Cráneo y mandíbula
de Mylodon exhibido en el Museo de
Antropología y Ciencias Naturales de Concordia,
Entre Ríos. |
Cueros, pelos y osteodermos en el Museo Argentino de
Ciencias Naturales de Buenos Aires. Seno de la
Ultima Esperanza. |
|
 |
 |
|
Excrementos de Mylodon
listai, del Seno de la Ultima
Esperanza (Patagonia Chilena) exhibidos Museo de La Plata. |
Recreación de
Mylodon (*) |
|
|
Esqueleto (*),
y
Por ultimo, una reconstrucción (*). |
Mamífero
Pilosa.
Perteneciente al infraorden
Pilosa,
Mylodon era de
menor tamańo que Megatherium
americanum, aunque compartía con él ciertas semejanzas
morfológicas; fue descubierto por primera vez por
Charles Darwin en
los alrededores de Bahía
Blanca y posteriormente estudiado por
Richard Owen, y se
alimentaba principalmente de vegetales. Se estima que alcanzaba unos
2,5 metros de altura a la
cruz y un peso aproximado de
tres toneladas. En
el Museo de la Universidad
Nacional de La Plata y en el
Museo Argentino de Ciencias
Naturales de Buenos Aires se exhibe un fragmento de
cuero momificado de Mylodon,
un hallazgo excepcional realizado de manera fortuita a fines del
siglo XIX en una caverna del
Seno de Última Esperanza, conocida como la
Cueva del Mylodon,
donde también se encontraron excrementos y grandes huesos dispersos
en sedimentos con restos de carbón, cuya reciente datación muestra
que estos restos de piel se momificaron hace unos 13.000 ańos atrás,
y que Mylodon convivió con los humanos. Este descubrimiento permitió
conocer la estructura de su piel, que contenía pequeńos huesecillos
subesféricos (osteodermos)
incrustados en el interior del cuero, los cuales le otorgaban
flexibilidad y, al mismo tiempo, una notable protección,
convirtiéndolo en un verdadero animal acorazado. Los paleontólogos
consideran que esta adaptación cutánea pudo haberse originado en sus
antecesores del Paleoceno
o incluso antes, y habría dado lugar posteriormente a los acorazados
dasipódidos y
gliptodóntidos.
En otros sitios de la Patagonia chilena y argentina se han
recuperado restos similares, aunque menos espectaculares, y un
hallazgo realizado en la localidad de
Tres Arroyos,
provincia de Buenos Aires, permitió comprobar que los milodontes
fueron cazados y consumidos, junto con otros grandes mamíferos, por
grupos humanos prehistóricos. Esta especie se extinguió entre
10.000 y 8.000 ańos antes
del presente, y no existe consenso entre los investigadores
respecto al número de taxones válidos del género, dentro del cual se
incluyen Mylodon darwini
Owen, 1839, registrado en la región pampeana de Argentina;
Mylodon listai Ameghino,
de menor tamańo, presente en la Patagonia esteparia argentina y
áreas similares de Chile; y
Mylodon insigne Kraglievich, también documentado en la región
pampeana argentina.
|
Lestodon
armatus.
Gervais, 1855. |
|
lestodon arnatus lestodon arnatus lestodon arnatus lestodon
arnatus estodon arnatus lestodon arnatus lestodon arnatus
lestodon |
 |
|
 |
|
Lestodonte.
Por
MarMag.2025. |
|
Esqueleto en el Museo de La Plata. |
|
 |
 |
 |
|
Cráneo de
Lestodon en el Museo de La Plata. |
Fémur de un
Lestodon que superaba a Megatherium |
Esqueleto completo de Lestodonte en el Museo Paleontológico
de Salto (Bs As). (*) |
Mamífero
Pilosa.
Fue un gran
xenartro
del Pleistoceno que, al igual que sus parientes, era principalmente
herbívoro y obtenía su alimento en las orillas de pantanos y cauces
fluviales, aunque posiblemente también actuaba como carrońero
oportunista. De gran tamańo, aunque menor que
Megatherium americanum, el
registro más antiguo del género se remonta al
Montehermosense
(Plioceno temprano); alcanzaba aproximadamente
4 metros de longitud,
con un cráneo de región rostral notablemente ensanchada, huesos de
los miembros grandes y robustos, y una dentición simple,
destacándose grandes dientes caniniformes proyectados hacia el
exterior de la boca, que habrían funcionado como defensa frente a
posibles depredadores. Su masa corporal se estima en unas
2,5 toneladas, y,
después de Scelidotherium,
Lestodon es uno de
los mamíferos fósiles del Cuaternario más frecuentes en el
territorio pampeano; junto con
Megatherium, representa a los
Tardigrada de
mayor tamańo corporal de su época. Investigadores del
Museo Paleontológico de
San Pedro dieron a conocer el descubrimiento de una manada
compuesta por diez individuos, tanto adultos como juveniles,
mientras que el Museo de
Ciencias Naturales de Miramar informó el hallazgo de al
menos cuatro ejemplares de distintas edades arrastrados por un río
hace miles de ańos, entre los cuales se recuperaron restos de un
individuo que habría superado en tamańo a
Megatherium, lo que abrió
nuevas hipótesis sobre su biología. Generalmente se considera que
estuvo adaptado a biomas abiertos de pastizales y arbustales, y
estudios tomográficos recientes realizados en los oídos de algunos
perezosos gigantes indican que eran animales sorprendentemente
rápidos y ágiles, comparables a hipopótamos o rinocerontes, en
marcado contraste con la visión tradicional que los describía como
criaturas extremadamente lentas. Entre las especies relacionadas se
reconocen Lestodon trigonidens,
exclusiva del Bonaerense y Lujanense, y
Lestodon armatus,
presente estratigráficamente en el Ensenadense y Lujanense, aunque
algunos autores sostienen que ambas podrían corresponder a una única
especie.
|
Scelidotherium leptocephalum.
Owen, 1840. |
|
Scelidotherium
Scelidotherium Scelidotherium Scelidotherium Scelidotherium
Scelidotherium Scelidotherium Scelidotherium
|
|
 |
|
 |
|
Esqueleto de Celidoterio (Scelidotherium
leptocephalum) exhibido en el Museo de La Plata. |
|
Esqueleto
montado por Fundación Azara en el Museo de
Ciencias Naturales de Miramar. |
|
|
|
 |
|
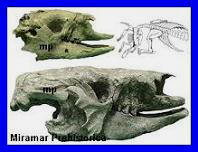
Cráneos de
Scelidotherium adulto y su cría, encontrados juntos.
Preservados en el Museo Cs Naturales de Miramar. |
|
Reconstrucción
de
Scelidotherium
en su cueva. Por Meredith. |
|
 |
|
 |
|
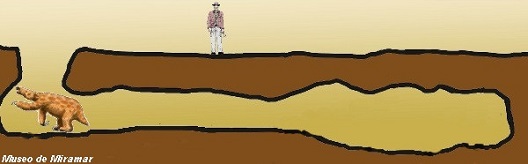
Una gran Paleocueva con
relleno y estructura interna, con una galería de 20 metros,
dos grandes cámaras y conexión a una antigua superficie, en
el Pleistoceno de Miramar, halladas por Mariano Magnussen
del Laboratorio Paleontológico del Museo de Ciencias
Naturales de Miramar. de las mismas se recuperaron marcas de
garras, atribuidas a Scelidotherium. |
|
|
|
Paleocueva de
Miramar. Una maravilla paleontológica bonaerense. En este esquema se dimensiona el recorrido
y tamańo total de la Paleocueva o Crotovina
realizada por un perezoso gigante y documentada por el Museo
de Miramar. Ilustración de D. Boh. |
Mamífero
Pilosa.
Los primeros restos fósiles
atribuidos a este género fueron descubiertos por
Charles Darwin
durante el viaje del Beagle
en la localidad de Bahía
Blanca, y a partir de materiales correspondientes al
cráneo, vértebras y costillas,
Richard Owen lo
denominó Scelidotherium en
1840. De menor tamańo que otros perezosos gigantes del Pleistoceno
mencionados previamente, estos
xenartros fueron,
sin embargo, algunos de los mamíferos más comunes del paisaje, como
lo demuestra la abundancia de sus restos en los depósitos
sedimentarios de este período; alcanzaban aproximadamente
1,7 metros de altura,
3,5 metros de longitud
y un peso estimado de una
tonelada. Su cráneo era alargado, bajo y estrecho, con un
rostro proyectado hacia adelante, y su dieta estaba compuesta por
vegetales obtenidos principalmente en ambientes secos. Estudios
biomecánicos realizados por investigadores del
Museo de La Plata
sobre los huesos de las extremidades anteriores demostraron que
Scelidotherium estaba
plenamente adaptado para la excavación, ya que la robustez de sus
huesos y sus amplias manos en forma de pala son coherentes con un
comportamiento de
superexcavador; este hábito fue corroborado a fines de la
década de 1990 con el descubrimiento de varias cuevas de estructura
interna cerca de la ruta 2, en las proximidades de
Mar del Plata, y
en 2002 con el hallazgo en
Miramar de una paleocueva con galerías, dos cámaras y
marcas de garras en paredes y techos, de unos
20 metros de extensión
y conectada con la antigua superficie. En ańos posteriores se
recuperaron esqueletos completos al sur de
Centinela del Mar
(Partido de General Alvarado), otro en
Camet Norte
(Partido de Mar Chiquita) y un notable ejemplar de una hembra adulta
abrazando a su posible cría en
Mar del Sud,
actualmente preservado en el
Museo de Ciencias
Naturales de Miramar. El género
Scelidotherium se
diferencia de Scelidodon
por la presencia de caracteres más derivados en los miembros y el
cráneo, y comprende al menos dos especies pleistocenas reconocidas,
Scelidotherium floweri y
Scelidotherium leptocephalum.
Estas especies habitaron ambientes abiertos y parcialmente arbolados
de climas templados, aunque durante el Pleistoceno tardío también se
registran en ambientes de altura, entre
2.000 y más de 3.000
metros, bajo condiciones templado-frías, como en
Las Juntas
(Catamarca) y Lauricocha
(Perú), si bien en este último sitio existen evidencias de que los
restos fueron transportados por el ser humano. Este notable mamífero
desapareció del registro fósil hace aproximadamente
7.000 ańos, y su
distribución geográfica abarcó los actuales territorios de
Argentina, Perú, Uruguay y
Paraguay; entre los géneros relacionados se incluyen
Proscelidodon patrius,
Scelidotherium floweri,
Scelidotherium parodi,
Scelidotherium leptocephalum,
Scelidodon patrius y
Scelidodon capellini.
|
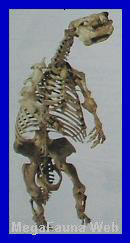
Glossotherium robustum.
Owen, 1842. |
|
glossotherium robustum glossotherium robustum glossotherium
robustum glossotherium robustum glossotherium robustum
|

Esqueleto adulto y de un juvenil en el Museo de La Plata.
(*) |
 |

Reconstrucción de Glosoterio. (*) |
Mamífero
Pilosa.
Fue un género extinto de
perezosos gigantes
sudamericanos, perteneciente a la familia
Mylodontidae,
caracterizado por su gran tamańo, hábitos completamente terrestres y
cuerpo corpulento, con una cabeza voluminosa y una cola larga y
pesada. Presentaba pies largos provistos de garras curvadas hacia el
interior, al igual que otros pilosos ya descritos, por lo que
caminaba apoyando los nudillos; es probable que pudiera erguirse
sobre las patas traseras, equilibrándose con la cola y utilizando
las garras para llevar el alimento a la boca. Alcanzaba
aproximadamente 3,5 metros
de longitud, 1,7
metros de altura y una masa estimada de
una tonelada, y su
extinción se produjo durante el
Holoceno medio,
hace unos 8.000 ańos antes
del presente. Su cuerpo estaba cubierto por una densa y
espesa pelambre, y en la piel se encontraban embebidos numerosos
osteodermos de
forma y tamańo variables, generalmente superiores a un centímetro de
diámetro. Los restos completos recuperados en Argentina son muy
escasos, y su dieta habría sido variada, compuesta por hojas de
gramíneas, arbustos, árboles y posiblemente carrońa. Los primeros
registros del género corresponden a la
Formación Vorohué,
y su desaparición habría estado vinculada a cambios climáticos
progresivos, sumados a la posible presión de la caza humana, que
redujeron paulatinamente sus poblaciones hasta la extinción. Un
género relacionado es
Paraglossotherium elmollarum, cuyos restos proceden de
Tafí del Valle,
en la provincia de Tucumán,
mientras que el registro más reciente de un ejemplar corresponde a
un fósil de unos 8.000
ańos de antigüedad, hallado a aproximadamente
160 km de la ciudad de
Buenos Aires. Estudios sobre la estructura del oído interno
indican que este animal era
rápido y ágil, en
marcado contraste con los perezosos arborícolas actuales. Otras
especies relacionadas incluyen
Glossotherium chapadmalense,
Glossotherium robustum y
Glossotherium myloides.
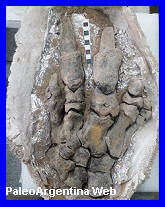
Catonyx tarijensis. Ameghino
1891.
|
 |
 |
 |
|
Cráneo de Catonix,
exhibido en el Museo de Ciencias Naturales de Miramar. |
Mano de Catonyx
tarijensis Río Tercero. Córdoba. |
Aspecto de
Catonyx tarijensis. Por Román Yevseyev. Tomado de Vk.com. |
Mamífero
Pilosa.
Es un género extinto de perezosos
gigantes terrestres perteneciente a la familia
Mylodontidae, endémico de
América del Sur
durante el Pleistoceno.
Fue nombrado por Florentino
Ameghino en 1891 y posteriormente asignado a la subfamilia
Scelidotheriinae
por Gaudin (1995).
Se trató de un gran herbívoro que probablemente compartía
preferencias ambientales y rasgos ecológicos con otros perezosos
gigantes de su época. Habitó el continente sudamericano desde hace
aproximadamente 2,5 millones
de ańos hasta cerca de
10.000 ańos antes del
presente, y sus restos fósiles han sido hallados en
Brasil y en la
Formación San José
de Uruguay; en
Argentina, en
cambio, el registro es escaso y se limita a algunos hallazgos en la
provincia de Córdoba
y en Miramar,
provincia de Buenos Aires.
Posiblemente, al igual que otros representantes de los
Pilosa, fue un
animal más ágil y rápido de lo que tradicionalmente se suponía, y
pudo haber incorporado
carrońa de manera ocasional a su dieta.
Scelidodon
copei. Ameghino, 1881.
|
 |
|
 |
|
Esqueleto de Scelidodon
en el Museo Argentino de Ciencias Naturales de Buenos
Aires.
|
|
Aspecto de Scelidodon.
Por Jorge Blanco. |
Mamífero Pilosa.
Se trata de un género extinto de perezosos
gigantes del Pleistoceno,
cuyos primeros restos fueron descriptos por
Charles Darwin en
Bahía Blanca
durante su viaje a bordo del
Beagle. A partir de cráneos, vértebras y costillas,
Richard Owen
interpretó que se trataba de un animal de gran masa corporal y
desplazamiento lento; sin embargo, descubrimientos posteriores
permitieron reconstruirlo casi por completo y confirmar un tamańo
considerable, comparable al de
Mylodon. Presentaba
un cráneo pequeńo, alargado, estrecho y bajo, con mandíbulas
prolongadas y premaxilares cortos, mientras que su dentición
mostraba superficies masticatorias elípticas o levemente
triangulares, con hileras dentarias paralelas y continuas de tamańo
similar en ambas mandíbulas, lo que indica una dieta estrictamente
herbívora basada en arbustos, hojas y raíces. Su cuerpo era robusto
y probablemente estaba cubierto por un espeso pelaje; alcanzaba unos
2,5 metros de longitud,
1 metro de altura
y un peso cercano a una
tonelada. Las extremidades posteriores eran largas y
poderosas, las anteriores más cortas, y ambas terminaban en grandes
garras curvadas hacia el interior, por lo que se desplazaba
apoyándose sobre el dorso de manos y pies; además, podía erguirse
sobre las patas traseras, utilizando su cola gruesa como soporte
para mantener una postura vertical. Sus registros más recientes
corresponden al
Pleistoceno tardío (edad Lujanense).
Pyramiodontherium
sp. Rovereto, 1914.
|
 |
|
 |
|
Fémur de Pyramiodontherium, recuperado
en el Pleistoceno de San Pedro, Buenos Aires. Imagen de
prensa. |
|
Posible
aspecto de
Pyramiodontherium. (*). |
Mamífero
Pilosa.
Los
Pyramiodontherium fueron un grupo de antiguos perezosos
terrestres que habitaron la Argentina desde finales del
Mioceno hasta el
Plioceno medio; sin
embargo, un reciente hallazgo realizado por el equipo del
Museo Paleontológico de San
Pedro amplía notablemente su rango temporal conocido. El
descubrimiento consiste en un
fémur prácticamente completo,
de 42 kilogramos de peso,
perteneciente a un individuo de tamańo colosal, lo que confirma la
presencia de este género hace apenas
700.000 ańos,
cuando hasta ahora se creía extinguido desde hacía unos
tres millones de ańos.
Este registro convierte al ejemplar de San Pedro en el más moderno
conocido para el género y sugiere que estos perezosos sobrevivieron
al menos dos millones de
ańos más de lo que se pensaba. Tanto el Grupo
Conservacionista de San Pedro como el Dr.
Luciano Brambilla,
investigador de la
Universidad Nacional de Rosario encargado del estudio del
material, coinciden en que las diferencias morfológicas observadas
en el fémur permitirían reconocer una
nueva especie,
hasta ahora desconocida dentro del género. Emparentados con los
megaterios, los
gigantescos perezosos que habitaron la región pampeana hasta fines
del Pleistoceno, los Pyramiodontherium eran considerados de tamańo
intermedio en comparación con aquellos; no obstante, la nueva
especie de San Pedro evidencia un desarrollo corporal excepcional,
que incluso supera al de los megaterios más grandes. Los restos de
este género son extremadamente escasos y se conocen únicamente en la
Argentina, con registros en las provincias de
Catamarca, La Rioja, Entre
Ríos y Chubut.
Neosclerocalyptus
ornatus.
Owen, 1845.
|
 |
 |
 |
|
Neosclerocalytus ornatus, exhibido en el
Museo Ciencias
Naturales en Villa Escolar, Formosa. |
Huellas fósiles de un
Gliptodonte del icnogenero
Glyptodontichnus
en el
sitio paleoicnologico de Puhuen Co. |
Neosclerocalyptus
ornatus, saliendo de su madriguera. Ilustración
de
Carlos Alberto Montefusco. |
Mamífero Cingulata.
Los
gliptodontes constituyen quizá la familia extinta de
mamíferos fósiles más conocida; su nombre, que significa “diente
tallado”, alude a la compleja morfología de su dentición. Su rasgo
distintivo era una coraza
rígida, sin bandas móviles como las de los armadillos
actuales, lo que limitaba su movilidad, formada por placas óseas
circulares e irregulares ensambladas como un mosaico. El origen del
grupo se remonta al Eoceno,
hace unos 45 millones de
ańos, aunque alcanzaron su máxima diversidad de formas y
tamańos hacia fines del
Plioceno y durante todo el
Pleistoceno. A
diferencia de sus parientes modernos, no eran insectívoros, sino que
se alimentaban principalmente de
pastos y vegetación dura.
Se conocen alrededor de 60
especies de gliptodóntidos, con tres morfologías básicas,
entre las cuales
Neosclerocalyptus representó una de las formas más pequeńas
de América del Sur. Este género presentaba un caparazón alargado y
bajo, con placas fácilmente identificables por el predominio de la
figura central sobre las periféricas; la cabeza estaba protegida por
un escudete óseo y la cola recubierta por anillos móviles y un tubo
largo y cilíndrico, posiblemente utilizado como defensa.
Neosclerocalyptus,
perteneciente a los
Hoplophorini, se caracteriza por el notable desarrollo y
neumatización de los senos frontonasales, la gran altura del cráneo
—especialmente en su región posterior— y la ausencia de cresta
sagital, reemplazada por una marcada concavidad media; sus huesos
nasales conferían al hocico un aspecto inflado, posiblemente
relacionado con el acondicionamiento del aire o con funciones de
exhibición. En la localidad de
Centinela del Mar,
cerca de Miramar, un equipo del Museo de La Plata identificó una
paleocueva con
estructura interna que contenía restos de este género, con marcas de
garras en las paredes y rastros de fricción del caparazón en el
techo, lo que, junto a ciertos rasgos anatómicos, sugiere posibles
hábitos cavadores
en estos gliptodontes de menor talla, cuyas extremidades eran
relativamente delgadas y cortas y cuyo peso oscilaba entre
250 y 300 kilogramos.
Sus restos son relativamente frecuentes en los sedimentos
cuaternarios del sudeste bonaerense, observándose diferencias entre
las especies más modernas, de unos
120.000 ańos, y
las más antiguas, que alcanzan hasta
un millón de ańos.
En 2024 se
publicó un estudio que analizó
32 marcas de corte
en fragmentos óseos de un ejemplar de
Neosclerocalyptus
hallado en la cuenca del
río Reconquista, en la localidad de Merlo, con una
antigüedad de 21.000 ańos,
demostrando una clara
interacción humana con la megafauna prehistórica; el patrón
de las marcas, interpretado como una secuencia lógica de desposte
con herramientas líticas, se suma a evidencias similares registradas
en América del Norte entre
26.000 y 22.000 ańos atrás. La distribución geográfica del
género es amplia e incluye
Argentina, Paraguay, Chile, Bolivia y Brasil, con un
registro que se extiende desde el
Ensenadense
(Pleistoceno temprano-medio) hasta el
Holoceno temprano,
asociándose principalmente a
ambientes abiertos de
pastizales parcialmente arbolados. Otras especies
relacionadas incluyen
Neosclerocalyptus heusseri, Neosclerocalyptus ornatus y
Hoplophractus.
Doedicurus clavicaudatus.
Owen, 1846.
Mamífero Cingulata:
Fue la
especie de mayor tamańo dentro de la familia de los gliptodontes,
alcanzando aproximadamente
4,3 metros de longitud y
2 metros de altura,
con una masa estimada cercana a
dos toneladas. Su
coraza presentaba una morfología particular, con una especie de
joroba dorsal en la
región más elevada y placas óseas unidas de manera más laxa que en
otros gliptodontes contemporáneos. La cola estaba conformada por
cuatro anillos,
seguidos por un tubo
cilíndrico de alrededor de un metro de longitud que
culminaba en una masa ósea
provista de púas córneas, constituyendo un eficaz sistema
defensivo frente a los depredadores habituales. Las extremidades
eran notablemente robustas, y tanto la columna vertebral como la
cadera se encontraban
fusionadas al caparazón, otorgándole una gran rigidez
estructural. Se habría alimentado principalmente de
pastos cortos en
ambientes semiáridos,
y, como el resto de los gliptodontes, se extinguió sin dejar
representantes actuales. Restos excepcionalmente bien preservados de
esta especie fueron recuperados durante
dos campańas
paleontológicas realizadas en 1918 por
Carlos Ameghino y Lorenzo
Parodi, y actualmente se encuentran depositados en el
Museo Argentino de
Ciencias Naturales “Bernardino Rivadavia” y en el
Instituto Nacional de
Investigación de las Ciencias Naturales, en Buenos Aires,
Argentina. Entre los géneros y especies relacionadas se incluyen
Neosclerocalyptus heusseri,
Sclerocalyptus ornatus, Hoplophractus, Plohophoroides conterminus,
Plohophorus figuratus, Panochthus tuberculatus, Doedicurus
clavicaudatus, Glyptodon munizi y
Paraglyptodon
chapadmalensis.
Glyptodon reticulatus.
Owen, 1845.
|
 |
|
 |
|
Extracción
de un Glyptodon reticulatus del Pleistoceno por el Museo
Municipal Punta Hermengo de Miramar. |
|
Ejemplar de Glyptodon munizi en el Museo de
Ciencias Naturales de Miramar. |
|
 |
|
 |
|
Excepcional
hallazgo de cuatro Gliptodontes, en el lecho del arroyo Salado, en el partido
bonaerense de Bolívar. |
|
Aspecto en vivo de
Glyptodon
reticulatus por Roman Uchytel. |
Mamífero Cingulata.
Fue una de las especies más comunes en el
territorio bonaerense y en gran parte de la Argentina, como lo
demuestra la abundancia de sus restos fósiles, y su existencia fue
comunicada por primera vez por
Richard
Owen en 1845. Este gliptodonte alcanzaba aproximadamente
3,5 metros de longitud
y 1,4 metros de altura,
con una masa estimada de 1,5
toneladas, y presentaba un cráneo protegido por una coraza
cefálica; como es característico del grupo, su caparazón carecía de
bandas móviles y medía cerca de
2,5 metros de largo por 2
metros de ancho, estando formado por placas óseas
engrosadas de entre 1 y 8
centímetros de diámetro, fácilmente reconocibles por su
patrón en forma de “flor” o “roseta”. A diferencia de otros géneros,
su cola no mostraba un tubo terminal anquilosado, sino que estaba
compuesta por siete
anillos móviles de diámetro decreciente, con placas de
morfología robusta y puntiaguda. Sus extremidades eran cortas y
fuertes, y gran parte de la columna vertebral y la pelvis se
encontraban fusionadas al
caparazón, otorgándole una estructura rígida; su dieta se
basaba principalmente en el
pastoreo de hierbas bajas
en ambientes abiertos, como pastizales y estepas, hipótesis
reforzada por estudios recientes del aparato hioideo.
Investigaciones publicadas en Francia por paleontólogos argentinos
sugieren además que varias especies de gliptodontes, incluido
Glyptodon, podían incorporarse sobre las patas traseras y
apoyarse en la cola, formando un
trípode para
sostener su pesado cuerpo. Su extinción ocurrió hacia
los 10.000 ańos antes del
presente, y dentro de los Glyptodontidae es el género con
mayor distribución
latitudinal, desde el sur de Argentina hasta Venezuela,
siendo dudosa su presencia en Centroamérica, especialmente en
México. El hallazgo de corazas y placas óseas de Glyptodon es
extremadamente frecuente en la región pampeana, y en
2019 se determinó
que Glyptodon clavipes
y Glyptodon reticulatus
corresponden a una misma especie, cuyas diferencias se debían a
variaciones ontogenéticas; posteriormente, en
2020, un equipo
del INCUAPA–CONICET (UNICEN)
recuperó restos de caparazones de
cuatro gliptodontes
en el lecho del arroyo Salado, en el partido de Bolívar. Entre los
taxones relacionados se incluyen
Paraglyptodon
chapadmalensis (Plioceno),
Glyptodon munizi, G.
principalis, G. laevis y G. gemmatus (Ensenadense), así
como G. perforatus, G.
elongatus y G. clavipes, presentes en el Bonaerense y
Lujanense.
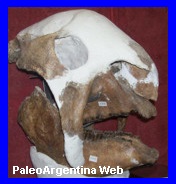
Panochthus
tuberculatus.
Owen,1845.
|
 |
 |
 |
|
Ejemplar de
Panochthus sp
en el Museo de La Plata. |
Cráneo en el
Museo de Cs Naturales de la localidad de Las Flores, Prov Bs
As. |
Panochthus, comparado con un armadillo
moderno del genero Dasypus. MarMag.2026. |
Mamifero Cingulata.
Fue otro de los célebres y
populares gliptodontes pampeanos, cuyo origen se remonta al
Eoceno,
a partir de pequeńas formas del tamańo de un gato doméstico, y que
alcanzó su máximo desarrollo durante el
Mioceno, Plioceno y,
especialmente, el Pleistoceno. Se diferencia de
Glyptodon por
su característica coraza en
forma de casco prusiano, formada por placas óseas gruesas
con un relieve compuesto por numerosas figuras circulares pequeńas,
uniformes y finamente punteadas; este caparazón presenta un marcado
alargamiento de la región
pélvica en detrimento de la zona torácica. El tubo caudal
era de gran tamańo y notablemente aplanado en comparación con otros
gliptodontes, y exhibe cicatrices que sugieren la presencia de
largas espinas córneas
puntiagudas, utilizadas como un eficaz sistema defensivo
frente a los carnívoros de la época. Su cráneo era corto pero
extremadamente robusto, acompańado por mandíbulas masivas y de
aspecto grotesco, mientras que las extremidades eran cortas y muy
gruesas; al igual que en otros gliptodontes, la
pelvis y gran parte de la
columna vertebral estaban fusionadas a la coraza dorsal,
otorgándole gran rigidez estructural. Los fragmentos de coraza de
Panochthus
son relativamente frecuentes en los depósitos pampeanos, y su
distribución geográfica abarcó los actuales territorios de
Argentina, sur de Brasil,
Uruguay y Bolivia, donde habría habitado ambientes abiertos
dominados por pastizales, de los cuales se alimentaba. Para el
Bonaerense y Lujanense
se reconocen varias especies, entre ellas
Panochthus tuberculatus,
P. morenoi y P. frenzelianus, que se
diferencian principalmente por el tamańo corporal y ciertos
caracteres craneales, como la presencia de órbitas abiertas o
cerradas, registrándose además el género
Panochthus intermedius.
|
Pampatherium typum.
Ameghino, 1937.
|
|
pampatherium typum pampatherium typum pampatherium typum
pampatherium typum pampatherium typum pampatherium |
|
Mandíbula de
Pampatherium en el Museo de La Plata, y coraza
articulada. Aspecto del armadillo gigante (*). |
Mamífero Cingulata.
La familia
Dasypodidae (o
Chlamyphoridae) agrupa a mamíferos cuyo cuerpo se encuentra
protegido por una coraza ósea formada por placas dérmicas, dividida
en una coraza escapular
y una coraza pélvica,
separadas entre sí por varias bandas móviles cuyo número varía según
la especie. En la actualidad, este grupo está representado por
animales de pequeńo tamańo, como los peludos y las mulitas; sin
embargo, durante el
Cuaternario existió una forma de gran tamańo conocida como
Pampatherium,
cuyo nombre significa “bestia de la pampa”. Este animal habría
alcanzado unos 225
kilogramos de peso y superado los
2,5 metros de longitud,
presentando un caparazón con bandas móviles que le permitían
realizar movimientos ágiles y excavar extensas galerías
subterráneas, donde probablemente pasaba gran parte de su tiempo.
Los registros más antiguos de los
Pampatheriidae en
la región pampeana, correspondientes al
Plio-Pleistoceno,
son relativamente escasos, aunque resultan más frecuentes en el
Mio-Plioceno de
la Mesopotamia, representados por el género
Kraglievichia, y
en la región sudandina, por
Vassalia; en ańos
recientes, diversos investigadores han vuelto a incluir a este grupo
dentro de la familia
Dasypodidae, tal como lo había propuesto en su momento
Florentino Ameghino. Se estima que su dieta era amplia y
oportunista, incluyendo gusanos, huevos, vegetales e incluso
carrońa, hipótesis apoyada por la morfología de sus dientes; en
particular, la presencia de una
cresta axial de dentina
compacta en los molariformes sugiere un consumo
predominante de vegetales
duros o altamente abrasivos, como las gramíneas.
Pampatherium typum
se registra en la Argentina a lo largo de todo el
Ensenadense y Lujanense,
con hallazgos en las provincias de
Buenos Aires, Santa Fe,
Córdoba, Corrientes y Formosa, y también fuera del país, en
la República Oriental del
Uruguay, Bolivia
(Tarija y Papúa) y Brasil.
Al igual que los gliptodóntidos, se lo asocia principalmente a
ambientes abiertos y
pastizales, y entre los géneros relacionados se reconocen
Plaina intermedia, Kraglievichia sp., Chlamydotherium paranense,
Pampatherium typum y Vassalia sp.
Propraopus grandis.
Ameghino, 1881.
|
Coraza y cola de
Propraopus grandis. Este ejemplar es el mas completo que se
conoce y fue hallado en el Pleistoceno bonaerense, exhibido en el
Museo de La Plata. Al lado, posible aspecto de Propraopus
grandis,
por MarMag.2025. |
Mamífero
Cingulata. Se trata de otro género de
dasipódidos, mamíferos
caracterizados por una coraza
ósea formada por placas córneas que, a diferencia de los
gliptodontes, presentaban bandas
móviles, lo que les permitía realizar amplios movimientos e
incluso enrollarse parcialmente en forma de bola. Su origen se remonta
al Paleoceno medio,
hace aproximadamente 55 millones
de ańos, y en un lapso relativamente corto lograron colonizar
una gran diversidad de ambientes, alcanzando una notable
diversificación. Su tamańo era comparable al del actual
tatú carreta que habita
el norte argentino, alcanzando cerca de
1,20 metros de longitud.
Su dieta era amplia y oportunista, compuesta principalmente por
gusanos, huevos, vegetales y
animales en estado de descomposición. Las extremidades eran
cortas y robustas, rasgo típico del grupo, con
falanges ungueales muy
desarrolladas, adaptadas para excavar extensas madrigueras
donde pasaban gran parte de su vida y donde las hembras parían a sus
crías. La presencia de
osteodermos fijos se reconoce por su forma hexagonal y por la
existencia de dos o tres figuritas periféricas en el borde anterior,
mientras que en las placas móviles se observan dos surcos que divergen
hacia la parte posterior, delimitando una figura subtriangular. El
registro fósil de este género es
muy escaso y suele
estar representado por restos aislados. Típicamente
pleistoceno, el
género incluye a P.
punctatus y P.
sulcatus en Brasil,
P. magnus en
Ecuador y P. grandis
en Argentina y Bolivia, con una distribución que se extiende hacia el
norte hasta Florida, Missouri
y Oklahoma, en los Estados Unidos. En la Argentina, la única
especie reconocida es P.
grandis, registrada en los pisos
Ensenadense y Lujanense
en las provincias de Buenos
Aires, Córdoba y Catamarca, y desde el punto de vista
paleoambiental su presencia sugiere
ambientes abiertos.
Chaetophractus villosus.
Desmarest, 1804.
|
 |
 |
 |
|
Ejemplar
mayormente completo de
Chaetophractus sp. Museo
Municipal de Ciencias Naturales de Miramar. |
Cráneo del mismo
ejemplar de la izquierda de
Chaetophractus sp. Museo
de Ciencias Naturales de Miramar. |
Aspecto
de
Chaetophractus.
Carlos Wiedner. |
Mamífero Cingulata.
Se trata de un
armadillo fósil emparentado con el género viviente conocido
como peludo pampeano,
cuyo linaje tiene un origen
sudamericano, con antecesores fósiles que se remontan al
Eoceno, hace
aproximadamente 45 millones de
ańos; posteriormente, tras el restablecimiento de la conexión
entre ambas Américas, este género logró expandirse hacia regiones del
centro de Norteamérica.
Era un armadillo fuertemente
acorazado, con una armadura compuesta por un
mosaico de pequeńas placas óseas
desarrolladas en la dermis y recubiertas por epidermis córnea, que
constituían un eficaz sistema de defensa contra los depredadores,
incluyendo la protección de la cola. Estas placas formaban un
escudo rígido sobre los
hombros y otro sobre los cuartos traseros, mientras que la región media
del cuerpo estaba cubierta por
bandas transversales articuladas, unidas por piel flexible, lo
que le permitía enrollarse
completamente y proteger el abdomen, la única zona vulnerable.
Presentaba un cuerpo robusto, con
patas cortas y musculosas
que le otorgaban gran agilidad, y extremidades provistas de
garras bien desarrolladas,
utilizadas tanto para la obtención de alimento como para la excavación
de madrigueras. De hábitos
nocturnos, su dieta era variada e incluía
insectos, gusanos, pequeńos
vertebrados y ocasionalmente carrońa. Los restos fósiles de
este y otros géneros morfológicamente afines son frecuentes en los
afloramientos sedimentarios del
Plioceno y Pleistoceno
pampeanos. Entre los géneros y especies relacionados se
incluyen
Chorobates recens,
Chorobates villosissimus,
Dasypus laevisculptus,
Macrochorobatus chapadmalensis,
Macroeuphractus outesi,
Macroeuphractus retusus
y
Zaedyus sp.
Lestodelphys halli.
Thomas, 1921.
|
 |
|
 |
|
Mandíbulas y maxilares
comparados con una moneda de cinco centavos para indicar
su tamańo real. |
|
Aspecto en
vida del pequeńo carnívoro marsupial Lestodelphys
del Pleistoceno Pampeano. (*) |
Mamífero Marsupial.
Se trata de un
género muy abundante durante el
Pleistoceno pampeano, caracterizado por su
pequeńísimo tamańo,
comparable al de un ratón, lo que habría hecho fácil confundirlo a
simple vista, aunque en realidad pertenecía a la
familia de las zarigüeyas o
falsas comadrejas (marsupiales). Habría presentado
hábitos tanto arborícolas como
terrestres, con una
dieta omnívora que incluía huevos, insectos y pequeńos roedores
contemporáneos de tamańo similar. Su
reproducción era compleja,
ya que las hembras parían crías muy poco desarrolladas que completaban
su crecimiento en un marsupio
externo. Sus restos fósiles son escasos y suelen encontrarse
asociados a otros
microvertebrados dentro de
regurgitaciones de aves rapaces,
lo que explica su frecuente fragmentación. Los géneros
Thylamys y
Lestodelphys son
los marmosinos fósiles más comunes en los registros
Plio-Pleistocenos de la región
pampeana, y sus representantes actuales muestran adaptaciones a
ambientes templados y xéricos,
como la capacidad de almacenar
grasa en la cola y de entrar en
estados de torpor diario o
hibernación, posiblemente como respuesta a la escasez
estacional de alimentos, especialmente insectos; es probable que sus
formas fósiles presentaran comportamientos similares. Como consecuencia
de los cambios climáticos y geográficos posteriores, su distribución
actual es más restringida, y hoy los representantes del género
Lestodelphys
habitan sectores de La Pampa y
el sur patagónico. Entre los géneros y especies relacionados se
incluyen Lestodelphys juga,
Lestodelphys halli,
Lutreolina crassicaudata,
Lutreolina tracheia,
Monodelphys dimidiata,
Thylamys contrerasi,
Thylamys pusillus,
Thylatheridium cristatum,
Thylatheridium pascuali,
Thylophorops perplana
y
Thylophorops chapadmalensis.
Lagostomus maximus.
Desmarest.1817.
 |
 |
 |
|
Cráneo con Mandíbula en el
Museo de Ciencias Naturales de Miramar. |
Restos muy
completos exhibidos en el Museo Paleontológico de San Pedro. |
Aspecto de Lagostomus maximus.
Por MarMag.2025. |
Mamífero
Roedor.
Se trata de una
especie de roedor
perteneciente a la familia Chinchillidae, representada en
la actualidad por la vizcacha (Lagostomus
maximus), la cual alcanzó una amplia expansión
geográfica recién a mediados del
Pleistoceno bonaerense.
Durante el Plioceno
existió un representante más antiguo y de menor tamańo,
Lagostomus (Lagostomopsis)
antiquus, mientras que el género es considerado
alóctono, ya que
habría llegado a Sudamérica durante el
Oligoceno tardío y el
Mioceno, posiblemente mediante procesos de dispersión como
el salto de islas, representado por formas tempranas como
Prolagostomus pusillus
y Lagostomus
telenkechanum. Su dieta estaba compuesta principalmente
por tubérculos, raíces y
hierbas, y presentaba un
cráneo ancho y robusto,
con nasales alargados,
bullas timpánicas poco
desarrolladas y mandíbulas provistas de una
cresta bien marcada.
Las extremidades
posteriores, más largas que las anteriores, poseían
tres dedos,
siendo el central el más desarrollado. Esta especie excavaba
extensas y complejas
madrigueras, con túneles y cámaras centrales, muchas de las
cuales pueden observarse en los afloramientos geológicos de
Miramar,
especialmente en las inmediaciones del
Bosque Vivero Dunícola
Florentino Ameghino, donde suelen hallarse restos fósiles
en su interior. En 1994,
Mariano Magnussen, del actual Laboratorio de Paleontología
del Museo de Ciencias Naturales de Miramar, Fundación Azara y LACEV,
descubrió en una de estas cámaras centrales restos de
cuatro individuos de
distintas edades, lo que constituye una clara evidencia de
comportamientos sociales
complejos en esta especie.
Hydrochoerus
sp. Brisson, 1762.
Roedor,
cavioideo.
En la actualidad, el
carpincho es el roedor más grande del mundo y pertenece al
suborden Caviomorphae,
familia Hydrochoeridae
y subfamilia Cavioidae,
aunque a lo largo de la historia geológica estuvo representado por
formas de tamańo aún mayor. La evolución de este roedor ha sido
ampliamente estudiada por paleontólogos uruguayos y argentinos, ya
que se trata de uno de los mamíferos que evolucionaron en América
durante la era Cenozoica,
con una filogenia que se remonta al
Oligoceno inferior
sudamericano. Durante el
Mioceno se produjo
una divergencia evolutiva entre los
Eocardidae e Hydrochoeridae
(capibaras primitivos), que evolucionaron de manera conjunta, y los
Cavioidae —grupo
que incluye a los cuises o cobayos—, cuyo desarrollo fue
independiente. En el
Pleistoceno existieron al menos
seis géneros con múltiples
especies, documentados por fósiles hallados desde el sur de
los Estados Unidos hasta el extremo austral de la Argentina,
correspondientes a individuos que alcanzaban hasta el
doble del tamańo de los
carpinchos actuales y mostraban una mayor especialización.
Las extremidades anteriores poseían
cuatro dedos y
las posteriores tres,
disposición similar a la del tapir (Tapirus
terrestris), con dedos unidos por
membranas natatorias
y provistos de uńas
fuertes, lo que indica una marcada adaptación a la vida
acuática, capaz de nadar con vigor y permanecer sumergido por
períodos prolongados, como ocurre hoy. Se han recuperado restos
fósiles relevantes en depósitos
Plio-Pleistocenos
de las provincias de Santa
Fe y Buenos Aires, y recientemente
Mariano Magnussen,
del Museo de Ciencias Naturales de Miramar, dio a conocer
huellas fósiles
atribuidas al icnogénero Porcellusignum
conculcator. De aspecto muy similar a sus parientes
actuales, estos roedores requerían
ambientes con abundante
agua dulce, lo que concuerda con el hallazgo de sus restos
en proximidad a antiguos
ríos, arroyos, lagos o pantanos, y su dieta habría estado
compuesta principalmente por
pastos y otras plantas.
A pesar de ello, el conocimiento sobre los carpinchos fósiles del
Pleistoceno aún es limitado, aunque sugiere una presencia
relativamente reciente, siendo
Neochoerus sp.
uno de los géneros extintos mejor conocidos para este período.
Desmodus draculae.
Morgan, Linares & Ray, 1988.
|
 |
 |
|
Rama
mandibular de Desmodus draculae, del Pleistoceno.
Museo de Ciencias Naturales de Miramar. |
Aspecto del
vampiro gigante del Pleistoceno, Desmodus draculae,
dentro de una paleocueva. Por HodariNundu. |
Quiroptero, desmodóntido.
fue una especie extinta de mamífero placentario del orden
Chiroptera,
que habitó regiones cálidas de
Centro y Sudamérica
durante el Pleistoceno
y el Holoceno reciente.
Los murciélagos vampiros son exclusivos del continente americano y
pertenecen a la familia
Desmodontidae, caracterizada por su dieta
hematófaga, siendo
el único grupo de murciélagos en el mundo con este hábito, rasgo que
históricamente despertó fascinación y dio origen a numerosas
leyendas. Se trata de animales de vuelo ágil y silencioso, con
notable capacidad para saltar, carentes de cola, de coloración
oscura y comportamiento generalmente hurańo y esquivo. En el
Cuaternario americano vivió
Desmodus draculae,
una forma estrechamente emparentada con el vampiro común, aunque
aproximadamente un 30 %
mayor en tamańo, descubierta por primera vez en
Venezuela en 1988.
En la Argentina, los únicos registros conocidos de vampiros fósiles
proceden del partido de
General Alvarado, donde se recuperó un canino superior
aislado del Holoceno
tardío en la localidad de
Centinela del Mar,
asignado a Desmodus cf. D.
draculae, cuya datación radiocarbónica arrojó una antigüedad de
apenas 300 ańos,
por lo que no llegó a fosilizarse completamente. Posteriormente, en
2021,
investigadores de la
Universidad Nacional de Mar del Plata, el
CONICET y el
Museo de Ciencias
Naturales de Santa Clara del Mar dieron a conocer el
hallazgo de una mandíbula
fósil de Desmodus draculae proveniente del
Pleistoceno del arroyo La
Ballenera, cercano a la ciudad de
Miramar,
actualmente depositada en las colecciones científicas del Museo de
Ciencias Naturales local. Este notable resto fue encontrado
in situ dentro de una
madriguera de 1,2 metros
de diámetro, atribuida a un
Mylodontidae, un
perezoso gigante, lo que abre diversas hipótesis, como la
posibilidad de que el vampiro se alimentara de estos animales,
utilizara la cueva como refugio grupal, o que el material haya sido
introducido por acción del agua o corresponda a restos de una presa.
Lo significativo es que tanto los registros fósiles como los
históricos de Desmodus
draculae se concentran en las cercanías de
Miramar, mientras
que su pariente viviente más próximo habita actualmente a unos
600 kilómetros al norte,
lo que sugiere un cambio ambiental y climático regional, siendo
probable que los últimos representantes de esta especie se hayan
extinguido durante la
época colonial, hacia comienzos del siglo XIX.
 |
El
gran Intercambio Biótico Americano (GIBA)
El
Gran Intercambio
Biótico Americano fue un suceso ocurrido durante
los últimos tres
millones de ańos, cuando se restableció la unión
terrestre entre
Sudamérica y Norteamérica, permitiendo el
desplazamiento masivo de fauna entre ambos hemisferios. Este
evento geológico posibilitó que numerosos mamíferos
sudamericanos migraran hacia el norte, como los
gliptodontes,
megaterios y toxodontes, mientras que desde el
hemisferio norte arribaron a Sudamérica
cérvidos,
mastodontes, équidos como Hippidion y diversos carnívoros,
incluidos osos y
tigres dientes de sable, entre otros. El impacto de
este intenso intercambio faunístico tuvo consecuencias
profundas y, en muchos casos, dramáticas, ya que generó una
fuerte competencia
por los nichos ecológicos, dificultades de
readaptación
ambiental y la introducción de
nuevas enfermedades,
factores que provocaron la disminución de muchas especies
autóctonas y llevaron a varias de ellas a la
extinción. |
Smilodon populator.
Lund, 1842.
|
Smilodon Smilodon
Smilodon Smilodon |
Smilodon Smilodon
Smilod |
Smilodon Smilodon
Smilodon Smilodon |
|

|

|
 |
|
Esqueleto de Smilodonte en el
Centro Cultural del Bicentenario, Santiago del Estero. |
Esqueleto de Smilodon en el
Museo de Miramar. |
Reconstrucción en vivo de
Smilodon. (*): |
|
 |
 |
 |
|
Cráneo y mandíbula
exhibida en el Museo Paleontológico de la ciudad de Mercedes (Bs
As). |
El técnico
Mariano Magnussen Saffer, con el calco de un cráneo del
Museo Municipal Punta Hermengo de la ciudad de Miramar. |
Primeras huellas atribuidas a Felipeda
miramarensis, un dientes de sable del Pleistoceno. Museo
de Miramar. |
Mamífero
Carnívoro.
El tigre dientes de sable fue uno de los
mayores logros evolutivos entre los mamíferos depredadores, y su
nombre, Smilodon, significa
precisamente “dientes de sable”, una característica que
refleja su extrema especialización para la caza de grandes presas,
como megaterios y mastodontes, hoy extintos. El primer hallazgo de
un smilodonte en la Argentina fue realizado por el naturalista
Francisco Javier Muńiz en 1844, en las barrancas del río Luján,
aunque el descubrimiento inicial del género ocurrió en 1842, cuando
el paleontólogo danés Peter Lund encontró restos en cavernas del sur
de Brasil y los atribuyó erróneamente a una hiena.
Smilodon superaba en
tamańo y peso al león actual, pero presentaba proporciones
corporales muy distintas a las de los félidos modernos: extremidades
posteriores cortas y robustas, cuello relativamente largo, lomo
corto y una poderosa región anterior del cuerpo. Su letalidad se
debía al extraordinario desarrollo de los músculos del cuello y los
hombros, junto con sus enormes caninos superiores, que podían
sobresalir más de quince centímetros; además, poseía una cola corta,
similar a la del lince. Sus mandíbulas podían abrirse en un ángulo
superior a los 120 grados, permitiendo que los largos dientes de
sable, de sección oval y aserrados en su borde posterior, se
hundieran profundamente en la carne de la presa con mínima
resistencia. Este formidable depredador habitó América durante más
de un millón de ańos, hasta que la llegada del ser humano y la
desaparición de la megafauna provocaron su extinción hace unos 8.000
ańos. En 2016, Mariano Magnussen, del Museo de Ciencias Naturales
Punta Hermengo de Miramar, descubrió en esa ciudad un sitio
paleoicnológico con huellas atribuidas a un gran tigre dientes de
sable, las cuales fueron estudiadas junto a a un equipoliderado por
el Dr Federico Agnolin, y presentadas en 2018 como una nueva
icnoespecie, Felipeda
miramarensis, única en el mundo.
Smilodon populator es la
especie de mayor tamańo entre todos los dientes de sable conocidos y
presenta dos subespecies:
Smilodon populator californicus, del Pleistoceno de América del
Norte, y Smilodon populator
populator, registrada en el Ensenadense y Lujanense de
Argentina, Brasil, Ecuador, Bolivia, Chile y Uruguay; un género
relacionado es Smilodon
fatalis, propio de Norteamérica.
Panthera onca.
Linneo,
1758.
|
 |
 |
|
Cráneo y
mandíbula de
Panthera de las barrancas del
río Bermejo de la localidad de General Mansilla,
recuperado por el Museo de Ciencias Naturales de Formosa. |
Panthera
onca atacando a un gran Gliptodonte. Por MarMag.2026. |
Mamífero
Carnívoro.
Es un carnívoro félido perteneciente a la subfamilia
Pantherinae, conocido vulgarmente como jaguar, yaguar o yaguareté,
siendo el mayor félido de América y el tercero más grande del mundo.
Su distribución actual se extiende desde el sur de Estados Unidos, a
través de América Central y gran parte de Sudamérica, hasta el norte
y noreste de Argentina. Durante el Cenozoico tardío, la familia
Felidae estuvo representada en la región pampeana por tres grandes
especies: Smilodon populator, Panthera onca
y Puma concolor, de las cuales S. populator se
extinguió a fines del Pleistoceno, mientras que P. concolor y
P. onca se registran con certeza desde el Ensenadense
(Pleistoceno temprano–medio), aunque en este último caso con
ejemplares aproximadamente un 30 % más grandes que los actuales.
Investigadores del CONICET, del Departamento Científico de
Arqueología de la Facultad de Ciencias Naturales y Museo de La Plata
y del INCUAPA, presentaron resultados recientes del sitio
arqueológico Nutria Mansa 1, en el partido de General Alvarado,
donde se halló un canino atribuido a Panthera onca; las dataciones
radiocarbónicas AMS realizadas sobre material dentario de Lama
guanicoe asociado permitieron vincular este registro al Holoceno
tardío en las inmediaciones de la costa atlántica pampeana, región
donde la especie se encuentra hoy localmente extinta. Asimismo, se
han reportado restos de otro cráneo en la localidad de San Miguel
de Tucumán. Aunque prefiere selvas densas y húmedas, el jaguar
puede adaptarse a una amplia variedad de ambientes boscosos o
abiertos, siempre asociado a la presencia de agua, destacándose
junto al tigre por su afinidad por la natación. Al igual que sus
representantes actuales, cazaba mediante emboscadas y era
oportunista en la selección de presas, desempeńando un rol clave en
la estabilidad de los ecosistemas. Los adultos poseían una mordedura
excepcionalmente potente, incluso en comparación con otros grandes
félidos, lo que les permitía perforar caparazones de reptiles
acorazados, como tortugas, y emplear un método de caza poco común,
atacando directamente la cabeza de la presa entre las orejas para
atravesar el cráneo con un mordisco fatal que alcanzaba el cerebro.
Panthera atrox.
Leidy,1853.
|
 |
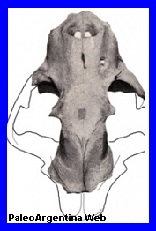 |
 |
|
Cráneo de Panthera atrox.
Muy similar al fósil preservado en el Museo de La Plata. |
Cráneo de Panthera atrox
recuperado en Patagonia. |
Aspecto de Pantera atrox
deambulando en la Patagonia Argentina. (*). |
Mamífero
Carnívoro.
El león americano fue aproximadamente un 25
% más grande que los leones africanos actuales (Panthera leo),
superando los 3 metros de longitud y los 250 kilogramos de peso, y
alcanzando un tamańo muy superior al de los yaguaretés. Se trataba
de una especie caminadora, adaptada a ambientes abiertos y secos,
como la estepa patagónica. A fines del siglo XIX, científicos
de distintas partes del mundo exploraron cuevas y cavernas de la
Patagonia con la esperanza de hallar con vida al Mylodon, un
perezoso gigante de hasta dos toneladas que, en realidad, se había
extinguido miles de ańos antes; aunque este objetivo no se concretó,
dichas expediciones permitieron el hallazgo de numerosos restos
fósiles, entre ellos esqueletos de un gran félido que durante
décadas fueron erróneamente clasificados. Santiago Roth, el primer
paleontólogo en estudiar y publicar estos restos, los consideró el
felino sudamericano más grande conocido y, al asociarlos con un
animal misterioso presente en las leyendas locales, los denominó
Lemish listai; posteriormente, se consensuó que
correspondían a una subespecie de yaguareté (Panthera onca)
de gran tamańo que habría habitado el extremo sur del continente.
Sin embargo, en diciembre de 2017, investigadores del Museo
Argentino de Ciencias Naturales “Bernardino Rivadavia” (MACN–CONICET)
reexaminaron los fósiles conservados en el Museo de La Plata y
concluyeron que pertenecen en realidad al león americano (Panthera
atrox), una especie considerada hasta entonces exclusivamente
norteamericana y estrechamente emparentada con el león de las
cavernas de Eurasia (Panthera spelaea), resultados que
fueron publicados en la revista Comptes Rendus Palevol. Las
expediciones arqueológicas en cavernas patagónicas también revelaron
grandes acumulaciones de huesos de Mylodon que, si bien inicialmente
se atribuyeron a la acción humana, presentaban marcas de dientes
agudos que evidencian la depredación por parte de un gran félido,
denominado por los investigadores como la “Pantera de la
Patagonia”. Asociados a los restos óseos de estos leones
americanos se hallaron fragmentos de cuero que indican que su pelaje
habría sido de tonalidad rojiza, rasgo coincidente con la
representación de un gran felino en una pintura rupestre descubierta
en la cueva de El Ceibo, en la provincia de Santa Cruz, durante la
década de 1970. Finalmente, Nicolás Chimento y Federico Agnolín,
autores del estudio, proponen revisar otros restos atribuidos a
supuestos “yaguaretés fósiles” para reforzar la evidencia de la
presencia habitual de este gran depredador en el sur de Sudamérica.
Leopardus geoffroyi.
d'Orbigny & Gervais, 1844.
|
 |
|
 |
|
Recreación in
situ de
Leopardus geoffroyi.
Museo Municipal Punta Hermengo de Miramar. |
|
Aspecto en
vida de
Leopardus geoffroyi. (*). |
Mamífero Carnívoro.
Es una especie de mamífero carnívoro perteneciente a la
familia Felidae, grupo que durante el Pleistoceno de Argentina
incluyó también a felinos de pequeńo tamańo, cuyos registros fósiles
son particularmente escasos. Algunos de estos taxones arribaron
desde América del Norte durante el Gran Intercambio Biótico
Americano, hace aproximadamente 3 millones de ańos, lo que dio lugar
a una notable diversidad conocida a partir de estudios actuales,
aunque aún pobremente representada en el registro paleontológico.
Los restos más antiguos corresponden a Felis vorohuensis,
datados en el Plioceno tardío–Pleistoceno temprano, mientras que
Felis geoffroyi se registra en el Pleistoceno superior y
Felis lujanensis (sinónimo de Lynchailurus colocolo)
en el Holoceno temprano, ambos en la provincia de Buenos Aires;
asimismo, se reconoce la presencia de Herpailurus en
Tierra del Fuego hacia el final del Pleistoceno–Holoceno.
Leopardus geoffroyi presentaba un tamańo similar o apenas
mayor al de un gato doméstico, con un peso habitual de entre 4 y 5
kilogramos, aunque podía alcanzar hasta 8 kilogramos, y se
alimentaba principalmente de roedores, pequeńos lagartos e insectos,
complementando ocasionalmente su dieta con ranas y peces, lo que lo
convertía en un depredador relevante dentro de su ecosistema. Entre
los félidos de mayor tamańo registrados para este período también se
encontraba Puma (=Felis) concolor.
Arctotherium latidens. Bravard
in Gervais y Ameghino, 1880.
|
|
|
|
|
 |
 |

|
|
Atlas o primera
vértebra cervical. Museo Municipal de Miramar. |
Arctotherium angustidens,
de la colección de la División Paleontología de Vertebrados
del Museo de La Plata.
Imagen de
proyectojuco. |
Reconstrucción
en vivo de
Arctotherium (*). |
|
 |
 |
 |
|
Arctotherium, que constituye el mayor exponente de la
familia Ursidae, recuperado en la ciudad de Buenos Aires en
el MACN. |
Cráneo y
mandíbula de Arctotherium juvenil, encontrado a la
altura de la localidad bonaerense de Junín.
Museo Legado del Salado.
|
El
prestigioso investigador Leopoldo
Soibelzon, del Museo de La Plata, compara el humero
del oso gigante
Arctotherium angustidens
con el humero de un elefante actual. Imagen de prensa. |
Mamífero
Carnívoro.
Fue un género extinto de mamíferos
carnívoros de la familia Ursidae, conocidos comúnmente como osos
sudamericanos u osos de cara corta, cuyos representantes
arribaron a América del Sur durante el Gran Intercambio Biótico
Americano, en un contexto en el que el continente carecía de
carnívoros placentarios como félidos, cánidos o úrsidos, lo que
favoreció una notable radiación adaptativa de estos inmigrantes
holárticos y la aparición de nuevos géneros y especies. En
Argentina, y particularmente en la región pampeana, se reconocen
cuatro especies del género Arctotherium, caracterizadas por
su gran tamańo, con masas corporales estimadas entre aproximadamente
300 y 1.200 kilogramos según la especie y el sexo, siendo la forma
del Ensenadense (Pleistoceno temprano–medio) la que
alcanzó las mayores dimensiones, mientras que las especies del
Bonaerense (Pleistoceno medio) y Lujanense (Pleistoceno
tardío–Holoceno temprano) presentaron tamańos relativamente menores.
Aunque la dieta de cada especie no ha podido establecerse con total
precisión, el análisis de la morfología dentaria sugiere que estos
osos predaban activamente sobre la diversa megafauna herbívora
pleistocénica, además de consumir carrońa, como lo evidencian las
lesiones observadas en sus dientes, y complementar su alimentación
con recursos ricos en hidratos de carbono, como frutas y miel,
indicados por la presencia de caries en numerosos restos fósiles. En
2011, el investigador Leopoldo Soibelzon, del Museo de La
Plata, presentó el ejemplar del mayor oso conocido que haya
habitado la Tierra, Arctotherium angustidens, el cual,
erguido, habría alcanzado unos tres metros de altura y superado la
tonelada y media de peso; sus restos habían sido descubiertos
durante la década de 1930, a 9,6 metros de profundidad, durante la
construcción del Hospital San Juan de Dios de La Plata, y
permanecieron durante décadas almacenados en el Museo de La Plata.
En Sudamérica, el registro fósil de Arctotherium se
extiende por Venezuela, Bolivia, Brasil, Uruguay, Chile y Argentina,
con una presencia documentada desde el Ensenadense (Pleistoceno
inferior–medio, hace unos 1,7 millones de ańos) hasta el Lujanense
superior (Pleistoceno tardío, hace aproximadamente 11.000 ańos).
Theriodictis platensis.
Mercerat, 1891.
|
Ejemplar de
Theriodictis platensis
en el Museo Paleontológico de San Pedro. Imágenes
tomadas de José Luís Aguilar. |
Mamífero
Carnívoro.
Fue un cánido de grandes dimensiones, cuyos ejemplares
adultos de Theriodictis
platensis alcanzaban entre 30 y 40 kilogramos de peso, que
arribó a América del Sur durante el Gran Intercambio Biótico
Americano y se registra en el Pleistoceno del continente, entre
aproximadamente 1,2 millones y 11.000 ańos antes del presente, con
especial presencia en la región pampeana. La especie fue descripta
originalmente por Mercerat en 1891 y posteriormente estudiada por
Kraglievich y Berman, reconociéndose numerosos ejemplares, entre los
cuales destacan restos procedentes de Santa Clara del Mar y
los individuos más completos hallados en sedimentos ensenadenses de
San Pedro y Ramallo, además de registros en una veintena de
localidades adicionales y en playas de Río Grande do Sul, en el
extremo sur de Brasil; recientemente,
Theriodictis ha sido
reconocido como un taxón característico y exclusivo del Ensenadense.
Los análisis paleoecológicos basados en índices morfométricos y
variables cualitativas indican que se trataba de un cánido
hipercarnívoro, adaptado a la persecución activa y captura de
mamíferos de mediano a gran porte, con masas corporales estimadas
entre 50 y 300 kilogramos, depredando principalmente cérvidos,
caballos, camélidos y pecaríes, y posiblemente también individuos
juveniles de especies de mayor tamańo. Estudios paleoecológicos de
la fauna correspondiente al tramo final del Lujanense vinculan la
extinción de varios mamíferos de más de 100 kilogramos con la
desaparición de este gran depredador, cuyos registros más antiguos
se remontan a aproximadamente 780.000 ańos, y que se encuentra
relacionado con otros cánidos fósiles como
Theriodictis tarijensis,
Canis gezi,
Cerdocyon ensenadensis,
Lycalopex proplatensis,
Dusicyon ameghinoi,
Canis nehringi y
Lycalopex avus.
Canis
gezi.
Kraglievich,
1928.
|
 |
 |

|
|
Cráneo y
mandíbula de
Canis nehringi.
(MACN), |
Cráneo y
mandíbula del canido Canis gezi (MACN), |
Aspecto
de Canis s.p en el Pleistoceno Argentino. (*) |
|
 |
|
 |
|
Recreación de
un ejemplar in situ de Canis nehringi en el Museo de
Ciencias Naturales de Miramar. Por M. Magnussen. |
|
Rama
mandibular de un juvenil de Canido indeterminado con sus
dientes y molares sin eclosionar, comparado con un ejemplar
adulto. Museo de Miramar. |
Mamífero
Carnívoro. Fue una especie de cánido
extinto de gran tamańo que habitó la Argentina durante el
Pleistoceno tardío y que es poco conocida debido a la escasez de su
registro fósil; los individuos habrían alcanzado un peso estimado de
entre 32 y 40 kilogramos. Estudios paleoecológicos de la fauna del
Ensenadense vinculan la extinción de varios mamíferos de más de 100
kilogramos con la desaparición de estos grandes cánidos
hipercarnívoros. Sus restos fósiles se restringen al centro-este
de América del Sur, con solo dos ejemplares recuperados en
sedimentos Ensenadenses de la región pampeana, en el centro de
Argentina, y un tercero hallado en el sur de Brasil. Los taxones
Canis gezi (Ensenadense)
y Canis nehringi (Lujanense)
comparten notables similitudes dentarias y craneanas asociadas a la
hipercarnivoría, lo que sugiere la existencia de un ancestro común
para ambos clados; además, un estudio de ADN publicado en 2010
relacionó directamente a
Canis nehringi con
Canis dirus,
proponiendo incluso que podría tratarse de un sinónimo. Entre los
géneros y especies relacionados se incluyen
Canis gezi,
Protocyon,
Speothos,
Theriodictis y
Chrysocyon
Lycalopex ensenadensis. F.
Ameghino, 1888.
|
 |
|
 |
|
Esqueleto de Lycalopex ensenadensis,
recreado para el Museo de Ciencias Naturales de
Miramar |
|
Posible aspecto de Lycalopex ensenadensis. (*): |
Mamífero Carnívoro. Es una especie extinta de
zorro del género Lycalopex
que habitó el centro-este del Cono Sur sudamericano durante el
Cuaternario y fue descripta originalmente en 1888 por el científico
argentino Florentino Ameghino bajo el nombre
Canis ensenadensis. El
holotipo fue exhumado en la Formación Ensenada, de edad
Pleistoceno temprano–medio, durante las obras de construcción
del puerto de la ciudad de La Plata, en un sector correspondiente a
la localidad de Ensenada, en el partido homónimo del nordeste de la
provincia de Buenos Aires. Los individuos de
Lycalopex ensenadensis,
también conocido como Cerdocyon
ensenadensis, habrían tenido un peso estimado de entre 4 y 6
kilogramos y una dieta omnívora, que incluía plantas, roedores,
frutas, ranas, lagartijas y aves. Este taxón se encuentra
estrechamente relacionado con otros cánidos sudamericanos como
Dusicyon culpaeus,
Dusicyon avus,
Dusicyon cultridens,
Dusicyon gymnocercus,
Lycalopex griseus y
Lycalopex inca.
Pteronura sp.
Gray, 1837.
|
 |
 |
|
Cráneo y
mandíbula de
Pteronura sp en el Museo Paleontológico de Diamante,
Entre Ríos. |
Aspecto de
Pteronura sp
del Pleistoceno. Por
Carlos C. Wiedner. |
Mamífero Carnívoro. Es un género de mamíferos
carnívoros de la familia Mustelidae, representado por la
nutria gigante Pteronura,
cuyos individuos alcanzaban aproximadamente 1,5 metros de longitud y
un peso cercano a los 45 kilogramos, mostrando un conjunto de
adaptaciones propias de un estilo de vida anfibio. Durante el
Pleistoceno, al igual que sus representantes actuales, habitaba
ríos y arroyos de agua dulce, ambientes que solían desbordarse en la
temporada de lluvias, y se alimentaba principalmente de peces, en
especial bagres, aunque también consumía cangrejos. Se considera que
el género extinto Satherium
habría sido el ancestro de la nutria gigante moderna, migrando hacia
Sudamérica durante el Plioceno o a comienzos del Pleistoceno. En la
actualidad, la nutria gigante habita América del Sur, región
donde también se distribuyen tres de las cuatro especies vivientes
del género Lontra.
Estudios de secuenciación de ADN realizados en 2005 sobre mustélidos
situaron la divergencia evolutiva de la nutria gigante entre
aproximadamente 5 y 11 millones de ańos atrás, y bajo esta nueva
evidencia el árbol filogenético propone que
Lontra fue el primer
linaje en separarse, seguido por
Pteronura, aunque los
rangos temporales de divergencia presentan cierto solapamiento.
Conepatus primaevus. Burmeister, 1864.
|
 |
|
 |
|
Cráneo y
Mandíbula de Conepatus primaevus, del Pleistoceno de la
localidad de Gobernador Castro. Museo de San Pedro. |
|
Posible
aspecto del genero Conepatus del Plio- Pleistoceno de la
región pampeana (*). |
Mamífero
Carnívoro. Los zorrinos conocidos de América
del Sur, tanto fósiles como actuales, son asignados al género
Conepatus,
perteneciente a la familia Mephitidae, integrada por carnívoros
pequeńos a medianos, de cuerpo delgado y alargado, extremidades muy
cortas y una cola bien desarrollada; muchas de sus especies poseen
glándulas situadas a ambos lados de la abertura anal que producen
secreciones de olor intenso y desagradable como mecanismo defensivo.
En sedimentos atribuibles al Plio-Pleistoceno temprano y
medio de las barrancas de la costa atlántica, en las cercanías de
Miramar, se registra la presencia del zorrino
Conepatus altiramus,
considerado posiblemente ancestral de las especies posteriores del
género, como Conepatus
primaevus, al que se vinculan los zorrinos vivientes de
Sudamérica. Tradicionalmente, los zorrinos fósiles y actuales fueron
incluidos dentro de la familia Mustelidae; sin embargo, estudios
filogenéticos recientes han demostrado que los Mephitidae
constituyen un grupo hermano de Mustelidae y Procyonidae, motivo
por el cual fueron elevados al rango de familia. Los registros más
antiguos de méfitidos en América del Sur corresponden al Ensenadense
(Pleistoceno temprano–medio), y los restos fósiles del género
Conepatus han sido
documentados en Venezuela, Perú, Bolivia, Brasil y Argentina.
Stegomastodon platensis. Ameghino,
1888.
|
|
|
|
|

|
 |

|
|
Esqueleto en el
Museo Argentino de Ciencias Naturales de Buenos Aires. |
Stegomastodon
en el Museo Rincón de Atacama, termas Rió Hondo. |
Cráneo de
Stegomastodonte en el Museo de Moreno. (*) |
|

|
 |

|
|
Reconstrucción
de Notiomastodon. (*). |
Esqueleto
recreado de
Stegomastodon platensis en el Centro Cultural del
Bicentenario, Santiago del Estero. |
Cráneo en el
Museo de La Plata. |
Mamífero
Proboscideo.
Fue un megamamífero que emigró desde América del
Norte durante el Gran Intercambio Biótico Americano y encontró en
América del Sur un nuevo territorio para expandirse; se trataba de
un proboscídeo, dotado de trompa, emparentado con los
mamuts extinguidos y los elefantes actuales. Era herbívoro y
alcanzó masas corporales de hasta 3,5 toneladas y alturas cercanas a
los 3,5 metros, habitando principalmente ambientes abiertos y
pantanosos, donde sus restos óseos quedaban incorporados al
sedimento tras su muerte, razón por la cual los hallazgos fósiles de
estos antiguos elefantes son relativamente escasos, aunque en ańos
recientes se recuperó un cráneo excepcionalmente completo en el
patio de una vivienda en la localidad bonaerense de Moreno. El
género Notiomastodon
—diferenciado en América del Norte desde, al menos, el Plioceno
temprano y presente allí hasta el Pleistoceno temprano–medio— está
representado en América del Sur por dos especies reconocidas:
Notiomastodon platensis
y Notiomastodon waringi.
La primera se caracteriza por defensas rectas, ligeramente curvadas
en su extremo anterior y recubiertas de esmalte, mientras que en la
segunda las defensas carecen de esmalte; además,
N. platensis presenta
una distribución más austral, abarcando Uruguay, Paraguay, Brasil y
gran parte de Argentina, con un biocrón que se extiende desde el
Ensenadense hasta el Lujanense (Pleistoceno temprano–tardío y
Holoceno temprano). En Argentina, sus restos han sido exhumados en
las provincias de Buenos Aires —donde se localiza el límite más
austral conocido para los proboscídeos, cerca de los 37° S—, Entre
Ríos, Córdoba, Santa Fe, Santiago del Estero y Corrientes, y se
infiere que este taxón mostraba una marcada preferencia por climas
templado-cálidos y hábitats abiertos o de sabanas arboladas,
alimentándose principalmente de vegetación arbustiva. Los
mastodontes sudamericanos son incluidos por los paleontólogos en dos
géneros principales,
Cuvieronius y
Notiomastodon, y aunque durante mucho tiempo en Argentina
solo se reconoció a
Notiomastodon platensis, la especie
Notiomastodon waringi,
conocida previamente en Brasil, Venezuela, Ecuador, Perú y
posiblemente Colombia, amplía el panorama de diversidad de estos
grandes proboscídeos extinguidos.
Hippidium
principale.
Lund, 1945.
|
|
|
|
|
 |
 |

|
|
Cráneo de
Hippidium, en el Museo de Ciencias
Naturales de Miramar. |
Esqueleto de
Hippidium
en el Museo Argentino de Ciencias Naturales de Buenos
Aires. |
Reconstrucción de caballo
americano extinguido. Tomado de Pinterest. |
Mamífero
Perissodactyla.
Fue un mamífero alóctono que arribó a
Sudamérica a comienzos del Pleistoceno durante el Gran Intercambio
Faunístico de fines del Plioceno, aunque teorías recientes de
algunos paleontólogos proponen que el género
Hippidion podría haberse
originado en Sudamérica y estar emparentado con
Pliohippus y
Dinohippus del Plioceno
de América del Norte. Se trataba de un caballo primitivo que se
extinguió antes de la llegada de los primeros colonos humanos, sin
dejar descendientes vivientes, y que posteriormente fue reemplazado
por el caballo europeo. Presentaba un aspecto similar al de las
cebras africanas, aunque de menor altura y constitución más robusta,
con extremidades cortas y anchas que le conferían una apariencia
maciza; poseía una cabeza proporcionalmente grande y una
característica protuberancia frontal reforzada por el hueso
nasal, además de cavidades delante de las órbitas que
posiblemente alojaban glándulas utilizadas para el marcaje
territorial, rasgo interpretado como indicativo de dimorfismo
sexual, motivo por el cual durante ańos fue clasificado bajo el
nombre Onohippidium.
Su peso pudo alcanzar los 400 kilogramos. El primer registro de un
caballo fósil sudamericano corresponde a un molar superior hallado
por Charles Darwin cerca de Bahía Blanca, identificado en 1840 por
Richard Owen, quien inicialmente lo asignó a
Equus caballus y
luego lo denominó Equus
curvidens. Desde entonces, se han recuperado numerosos
esqueletos bien preservados en distintas regiones del país, entre
ellas Tafí del Valle, en la provincia de Tucumán, y su registro
fósil se extiende ampliamente desde el norte de Ecuador hasta el sur
patagónico. Estratigráficamente,
Hippidion es conocido
desde el Plioceno superior (Uquiense) hasta el Pleistoceno
superior–Holoceno (Lujanense reciente), y las adaptaciones
morfológicas de la porción distal de sus extremidades sugieren una
preferencia por ambientes de bosque húmedo y suelos blandos. Los
équidos sudamericanos se extinguieron por completo, aunque algunos
de los registros más recientes de
Hippidion provienen
de sitios arqueológicos de la provincia de Buenos Aires, como Arroyo
Seco, en el partido de Tres Arroyos, con una antigüedad aproximada
de 8.500 ańos. El género
Hippidion está representado por tres especies:
Hippidion devillei,
Hippidion principale
y Hippidion saldiasi,
las dos primeras registradas en la provincia de Buenos Aires y en
Tarija (Bolivia), y la última característica de la Patagonia
austral.
Tapirus dupuy.
C. Ameghino, 1916.
|
 |
|
 |
|
Cráneo de
Tapirus sp, recreado in situ. Museo de Ciencias
Naturales de Miramar. |
|
Aspecto en
vida de Tapirus. Por
Carlos C. Wiedner.
|
Mamífero
Perissodactyla.
Es una especie extinta de mamífero perisodáctilo de la familia
Tapiridae, perteneciente al género
Tapirus, que
habitó América del Sur durante el Pleistoceno y fue descripta
originalmente por Carlos Ameghino en 1916 a partir de materiales
fragmentarios recolectados en sedimentos del Ensenadense de la
ciudad de Miramar, provincia de Buenos Aires, en homenaje a José
María Dupuy (h), prefecto local y fundador del primer museo de
ciencias naturales de esa ciudad en 1912. Un estudio integral de los
materiales sudamericanos asignados al género
Tapirus concluyó que
Tapirus rioplatensis
constituye una especie dudosa debido a la escasez y fragmentación
del registro fósil, situación similar a la de
Tapirus australis (Rusconi,
1928). Al igual que otros integrantes del género, este taxón se
asocia a climas cálidos y ambientes de selvas lluviosas, sabanas o
bosques húmedos tropicales y subtropicales, generalmente próximos a
ríos o zonas pantanosas, con una dieta herbívora basada en hojas,
frutos y otros vegetales. Se trataba de un animal de tamańo mediano
a grande, con una longitud estimada entre 1,8 y 2,5 metros y un peso
de aproximadamente 220 a 300 kilogramos, caracterizado por su hocico
alargado en forma de pequeńa probóscide, utilizado para arrancar
hojas, hierbas y raíces. Los análisis morfométricos dentarios
indican que Tapirus
rioplatensis habría sido significativamente mayor que
Tapirus terrestris,
Tapirus mesopotamicus
y Tapirus rondoniensis,
y ligeramente más grande que otros tapires fósiles o actuales de
gran tamańo, como Tapirus
oliverasi, Tapirus
indicus, Tapirus
tarijensis y
Tapirus haysii.
Hemiauchenia paradoxa.
Gervais y Ameghino, 1880.
Mamífero
Artiodactyla.
Durante el Lujanense, las llamas y los guanacos no se encontraban
restringidos a su actual distribución andino-patagónica, sino que
eran habitantes frecuentes de las praderas pampeanas; estos
camélidos, originarios de América del Norte, arribaron al sur del
continente hace más de 1,5 millones de ańos. El registro más antiguo
de la familia Camelidae en la región corresponde al Plioceno
de Barranca de los Lobos, entre Mar del Plata y Miramar (provincia
de Buenos Aires), en la Formación San Andrés (edad Uquiense), y para
el Pleistoceno ya se encontraban bien diferenciados géneros como
Hemiauchenia,
Palaeolama,
Eulamaops y
Lama. Su apariencia
general era similar a la de las llamas y guanacos actuales,
aunque algunas especies alcanzaban tamańos excepcionales, con
alturas estimadas de hasta 2,5 metros, superiores incluso a las de
los camellos vivientes de Asia y África, y masas corporales
cercanas a una tonelada; eran principalmente pastadores y sus restos
son muy abundantes en la provincia de Buenos Aires hasta hace unos
8.000 ańos, momento a partir del cual fueron reemplazados por los
géneros vivientes, que tras el siglo XVII desaparecieron
completamente de la región pampeana, sobreviviendo únicamente en las
estepas patagónicas.
Palaeolama y
Hemiauchenia son especialmente frecuentes en los sedimentos
pleistocenos y se caracterizan por poseer miembros robustos,
metapodios cortos y mandíbulas mesognatas y bajas;
Palaeolama weddelli,
estudiada por Gervais en 1855, fue un camélido de gran tamańo, hasta
un tercio mayor que el guanaco actual, con un cráneo alargado y
hocico delgado, y se asocia principalmente a áreas andinas. También
se registra Eulamaops
parallelus, descripta por Florentino Ameghino en 1884 a
partir de escasos restos recolectados por su hermano Carlos en las
barrancas del río Luján, mientras que
Lama guanicoe, de menor
tamańo, corresponde al guanaco actual, con un linaje fósil conocido
desde el Pleistoceno a través de
Lama gracilis. Los
restos de Hemiauchenia
se han hallado ampliamente en Sudamérica, desde Bolivia hasta el
sur de la Patagonia, incluyendo Chile, Uruguay y gran parte de
Argentina, con abundancia notable en las pampas, así como en sitios
como Río Hondo (Santiago del Estero) y Tafí Viejo (Tucumán),
y se reconocen como géneros relacionados a
Lama lama,
Lama vicugna y
Lama owenii.
Antifer cf.
ensenadensis.
Ameghino, 1889.
|
 |
 |
 |
|
Fragmento de cornamenta de Antifer.
Depositada en el Museo de Ciencias Naturales Punta Hermengo de
Miramar. |
Vitrina del Museo de La Plata con restos de cervidos. En este
caso ilustrativo, el genero Morenelaphus. |
Reconstrucción del gigantesco cervido Antifer.
Por
MarMag.2025.. |
Mamífero
Artiodactyla.
La familia Cervidae ingresó a América del Sur durante el Gran
Intercambio Biótico Americano, en el límite Plio–Pleistoceno, y
experimentó una rápida radiación adaptativa a lo largo del
Pleistoceno, alcanzando una notable diversificación hacia fines del
Piso Ensenadense (Plioceno tardío–Pleistoceno medio),
evidenciada por la presencia de numerosos géneros tanto extintos
como actuales. La mayor parte de los registros procede de la región
Pampeana, cuyas especies resultan comparables con las faunas de
ciervos pleistocénicos de Río Grande do Sul, en Brasil, y de la
Formación Sopas (Pleistoceno superior) de Uruguay; en este contexto,
el material atribuido a
Antifer cf. ensenadensis es exclusivo del Ensenadense y se
reconoce tanto en la región Pampeana como en la provincia de
Santa Fe. Antifer
mostraba preferencia por ambientes abiertos con predominio de
vegetación arbustiva y presentaba una marcada especialización
ramoneadora, lo que sugiere que las condiciones ambientales del
Bonaerense y Lujanense de la Mesopotamia argentina habrían sido
similares a las del sur de Brasil y parte de Uruguay, caracterizadas
por climas algo más húmedos que los de la región Pampeana, la cual
habría mantenido condiciones relativamente más frías y áridas. Esta
interpretación se ve respaldada por evidencias sedimentológicas,
palinológicas y paleofaunísticas que indican una fuerte similitud
climática regional durante ciertos intervalos del Pleistoceno.
Morfológicamente, Antifer
se distinguía por poseer astas robustas y largas, de hasta 60
centímetros, comprimidas lateralmente, levemente arqueadas y con una
ramificación dicotómica irregular, cuya superficie exhibía surcos
longitudinales. Entre los géneros relacionados se incluyen
Blastocerus dichotomus,
Epieuryceros,
Morenelaphus,
Hippocamelus,
Paraceros y
Ozotoceros bezoarticus.
Morenelaphus brachiceros.
Carette, 1922.
 |
|
 |
 |
|
Esqueleto en el Museo
Provincial de Ciencias Naturales
de Córdoba. Restos de
cráneo hallado en Centinela del Mar, depositado en
el Museo de Ciencias Naturales de Miramar. |
Aspecto que presentaba en vida
Morenelaphus.
Por MarMag.2025. |
|
Mamífero
Artiodactyla.
Es otra de las especies que arribaron a comienzos del Pleistoceno.
Pertenece a la familia Cervidae, aunque corresponde a un género
totalmente extinguido y de mayor tamańo que los representantes actuales
del grupo. Poseía cuernos muy robustos, cilíndricos y parcialmente
achatados, arqueados longitudinalmente y con forma de “S”, rematados en
tres puntas muy filosas, ideales para la defensa. Su alimentación y
comportamiento no habrían diferido sustancialmente de los de los ciervos
modernos. Hallazgos muy relevantes de estos cérvidos fueron realizados
en territorio bonaerense por Carlos Ameghino y Jurado en 1913, por
Lorenzo Parodi en 1914 y por Lucas Kraglievich en 1928. En la
actualidad, los descubrimientos de estos mamíferos son abundantes en la
localidad fosilífera de Centinela del Mar, a unos 45 kilómetros de la
ciudad de Miramar, donde se conserva un enorme médano “fósil” con una
antigüedad estimada de 100.000 ańos antes del presente. Su presencia en
el registro estratigráfico se remonta al Bonaerense y se extiende hasta
el Lujanense tardío. En América del Sur, sus restos han sido hallados en
Uruguay, Paraguay, el sur de Brasil y Argentina. Existe, sin embargo,
escasa información sobre los requerimientos ecológicos de este género,
aunque algunos autores proponen ambientes abiertos o parcialmente
arbolados. Otras especies relacionadas incluyen Morenelaphus
lujanensis, Blastocerus dichotomus, Epieuryceros, Hippocamelus,
Paraceros y Ozotoceros bezoarticus.
Calomys musculinus.
Thomas, 1913.
|
 |
|
 |
|
Hemimandibula
de Calomys del Pleistoceno de Miramar. Museo de Ciencias
Naturales Punta Hermengo. |
|
Aspecto de
Calomys sp (*). |
Roedor, Cricetido.
Se
encuentran entre los vertebrados más pequeńos registrados en el
Pleistoceno. Se han recuperado fósiles del género Calomys
correspondientes al Ensenadense, Bonaerense y Lujanense, es
decir, de los últimos 700.000 ańos antes del presente. Los registros
más antiguos de cricétidos sudamericanos datan del Plioceno
temprano. Estos roedores son de origen norteamericano y aún se
debate el momento exacto de su ingreso a Sudamérica. Habrían pesado
alrededor de 20 gramos y su dieta incluiría granos, flores y
pequeńas ramillas. Habitaban cuevas o huecos naturales entre la
vegetación. Es una especie frecuente en el registro fósil dentro de
egagrópilas (bolos alimenticios) regurgitadas por aves rapaces,
donde suele aparecer asociada a numerosas especies de vertebrados
contemporáneos. Otros géneros conocidos son Akodon,
Reithrodon, Tafimys, Phyllotis, Neotomys,
Andinomys, Oligoryzomys, Oxymycterus,
Necromys, Abrothrix y Oryzomys.
Eubalaena sp.
Demoulins, 1822.
|
 |
|
|
 |
|
Humero atribuido tentativamente a un ejemplar
adulto de
Eubalaena sp,
Museo de Ciencias Naturales de Miramar. |
|
Mamífero
Cetáceo.
Representante fósil de la actual ballena franca, presentaba un cráneo
ancho y posteriormente deprimido. Poseía una columna vertebral larga,
compuesta por 58 o 59 vértebras: 7 cervicales, 15 dorsales, 10 lumbares
y 27 caudales, además de 15 pares de costillas, un húmero macizo de
longitud similar al cúbito y al radio, y cinco dedos cortos, con
excepción del medio, notablemente más largo. Al igual que sus
representantes actuales, se alimentaba de pequeńos crustáceos y
microorganismos que filtraba mediante las barbas ubicadas en la
mandíbula superior. En una zona terrestre de la localidad de Mar del Sud,
a unos 17 kilómetros al sur de la ciudad de Miramar (provincia de Buenos
Aires), se hallaron restos fósiles de una ballena del género
Eubalaena, datados en cientos de miles de ańos. Se recuperaron
principalmente huesos de la región posterior del cráneo, partes próximas
al oído y la articulación mandibular, lo que permitió un diagnóstico
confiable y puso en evidencia una particularidad: los fósiles de
ballenas del Cuaternario son extremadamente escasos. En algunas regiones
del país se conocen restos aislados de este género, sobre todo en las
cercanías de La Plata. La rareza de estos hallazgos se debe a que las
condiciones climáticas y marítimas favorecen el deterioro de los restos,
dificultando su preservación, sumado a que es excepcional encontrar
fósiles de origen marino en contextos continentales. La explicación
radica en las variaciones del nivel del mar a lo largo de los milenios.
Durante los períodos glaciares, el borde costero de la región pampeana
se encontraba hasta 200 kilómetros más alejado que en la actualidad, ya
que grandes volúmenes de agua quedaban retenidos en los glaciares. En
los períodos interglaciares ocurrió el fenómeno opuesto, con ascensos
del nivel del mar de hasta seis metros sobre el nivel actual, cubriendo
áreas hoy emergidas. Este animal habría muerto durante una de estas
fases, denominada “ingresión marina”, quedando sepultado en condiciones
favorables para su fosilización. Cabe destacar que a lo largo de la
historia se registraron varias ingresiones, entre ellas una ocurrida
hace 120.000 ańos y otra, más reciente, responsable de la formación de
la laguna Mar Chiquita hace unos 6.000 ańos (Formación Querandí). Por la
coloración del fósil y ciertas características particulares, se presume
que los restos pertenecerían a la ingresión más antigua, conocida como
“Belgranense continental”. Otras especies relacionadas incluyen
Plesiocetus garopii, Megaptera longipinna y
Eubalaena australis.
Notiocetus romerianus.
Ameghino, 1891.
 |
|
 |
|
Bula
timpánica de un Balaenopteridae |
|
Aspecto de la
ballena
Balaenoptera.
Tomada de soulpix. |
Mamífero Cetáceo.
Esta especie se conoce a partir de material muy fragmentario,
específicamente una bula timpánica aislada, publicada por Florentino
Ameghino. Sin embargo, estudios futuros y el hallazgo de nuevos
materiales podrían aportar conclusiones diferentes. Los cetáceos
Balaenopteridae están ampliamente representados en el registro fósil
del Mioceno temprano de Argentina, pero son sumamente escasos
durante el Pleistoceno, con excepción de unas pocas muestras sin
estudios detallados. La familia Balaenopteridae se distingue de
otras familias de misticetos por la presencia de pliegues en la
garganta y la región ventral (pliegues gulares), que se expanden
notablemente durante la alimentación al tragar grandes volúmenes de
agua que luego es filtrada por las barbas. Presentan una aleta
dorsal y el cráneo, visto lateralmente, es recto y plano.
Pseudoplatystoma.
Bleeker, 1862.
|
 |
|
 |
|
Porción
anterior del techo del cráneo (neurocráneo) de un
Siliriforme, hallado en las barrancas del río
Carcarańá, Santa Fe. Prensa. |
|
Aspecto de un
Siliriforme del genero Pseudoplatystoma. (*) |
Pez,
Siluriformes.
Conocidos comúnmente como peces gato, son endémicos de América del
Sur. Un equipo de investigadores que relevaba activamente las
barrancas del río Carcarańá en busca de fósiles del Pleistoceno
tardío halló restos de un cráneo. Estas tareas, realizadas mediante
la navegación paciente y metódica del río, permiten detectar fósiles
directamente desde el agua, facilitando su localización y
recuperación. En los sedimentos de la zona, al igual que en el resto
de la región pampeana, los restos de peces son extremadamente
escasos, ya que suelen poseer huesos cartilaginosos o de baja
densidad, lo que dificulta su preservación. Este fósil de surubsdo
fue hallado en un tramo del río Carcarańá que atraviesa el distrito
de Oliveros, provincia de Santa Fe, cerca de su desembocadura en el
río Paraná. El material corresponde a la porción anterior del techo
del cráneo (neurocráneo), con una longitud de 17 cm y un ancho de 6
cm, y se infiere que el animal habría alcanzado un tamańo similar al
de un surubí actual de entre 10 y 15 kg. El equipo del Museo
Paleontológico de San Pedro ha registrado también peces de diversas
familias que aún habitan las aguas del Paraná. Los relevamientos
permitieron recuperar restos de las familias Characidae (dorados),
Doradidae (armados) y Pimelodidae (bagres). Algunos de estos peces,
que pueden alcanzar hasta 100 kg de peso, se alimentan de peces
jóvenes de diversas especies, como sábalos, bogas y morenas, e
incluso de anfibios y reptiles, como ranas y pequeńas serpientes.
Oligosarcus
sp. Günther,
1864.
|
 |
|
 |
|
Parte del
cráneo atribuido a Oligosarcus
sp de las colecciones del Museo de Cs Naturales de
Miramar. |
|
Aspecto en
vida que presentaría Oligosarcus. |
Pez,
Characiforme.
Es un
género de peces conocidos comúnmente como dientudos o dentudos,
perteneciente a la familia Characidae y al orden Characiformes. La
ubicación filogenética del género dentro de la familia ha sufrido
numerosas modificaciones en la historia reciente de la sistemática
de los characiformes. En la actualidad, Oligosarcus se
distribuye en los ríos costeros del sudeste de Brasil, el sistema
lagunar costero de Río Grande do Sul, Uruguay, la Cuenca del Plata y
los cursos de vertiente atlántica de la provincia de Buenos Aires,
hasta Bahía Blanca. Habita tanto ambientes lóticos como leníticos y
presenta hábitos carnívoros, alimentándose principalmente de
insectos y crustáceos y, en menor medida, de otros peces. Como
ocurre con la mayoría de los peces cuaternarios, los restos fósiles
de este género son muy escasos. Los primeros registros fósiles de
Oligosarcus provienen de facies lacustres del Pleistoceno
medio de la localidad de Centinela del Mar, cercana a la ciudad de
Miramar. El material consiste en huesos dentarios aislados con una
combinación de caracteres única, que permite una identificación
segura a nivel genérico. Este hallazgo constituye el primer
characiforme fósil descripto para la región pampeana argentina.
Recientemente, Daniel Tassara, del Museo de Ciencias Naturales
Pachamama de Santa Clara del Mar, aportó restos craneanos
encontrados muy cerca del ejemplar original, los cuales fueron
entregados al Museo de Ciencias Naturales Punta Hermengo de Miramar
para su estudio y exhibición. Este pez, sin dudas, habría nadado
esquivando las enormes patas de perezosos gigantes y toxodontes que
se acercaban a las antiguas lagunas bonaerenses.
Synbranchus marmoratus. Bloch, 1795.
|
.jpg) |
|
.jpg) |
|
Vértebra de
Synbranchus sp, del
Pleistoceno de la localidad bonaerense de San Pedro.
|
|
Ilustración de
Synbranchus sp,
realizada por el PaleoArtista Miel guel ÁngLugo.
|
Pez. El
conocimiento de esta especie, está constituida por algunas vértebras
caudales asignadas tentativamente a este género y especie. Las
mismas, proceden de la Formación Río Bermejo, de la localidad de
Mansilla, en la Provincia de Formosa, los cuales, durante la época
seca y fría del Último Máximo Glacial. La paleomastofauna exhumada
asociadas a los restos de Synbranchus, son
mamíferos típicos de la edad Lujanense, como Toxodon,
Glyptodon, Scelidotherium, Hemiauchenia, entre otros, que
son una asignables al Pleistoceno tardío-Holoceno temprano,
sugiriendo la presencia de ambientes abiertos y áridos/semiáridos.
En el Río Quequén Salado (Provincia de Buenos Aires, Argentina) se
han encontrado fósiles de Synbranchus (específicamente
Synbranchus sp.) que datan del Pleistoceno
Superior. Estos fósiles son parte de una asociación ictiofaunística
que incluye otros 7 taxones distintos, incluyendo un "morfotipo
Quequén Salado" de Corydoradinae. Si bien se han encontrado fósiles
de Synbranchus en otros sitios pleistocenos de la
provincia, los fósiles del Río Quequén Salado son relativamente
escasos. Por el tamańo de las vértebras encontradas, se estima que
estos Synbranchus podían alcanzar una longitud de 1.20
metros. Así mismo, otra vértebra aislada procede del Pleistoceno
tardío de la localidad de San Pedro, en la Provincia de Buenos
Aires. En la actualidad, la Anguila Criolla (Synbranchus
marmoratus). Este pez de agua dulce se encuentra
principalmente en América del Sur y Central, donde habita en ríos,
pantanos y áreas inundadas. Synbranchus marmoratus ha
evolucionado a lo largo de millones de ańos para adaptarse a los
diversos hábitats acuáticos de América del Sur. A lo largo de su
historia evolutiva, este pez ha desarrollado características que le
permiten sobrevivir en condiciones ambientales cambiantes, como la
capacidad de respirar aire atmosférico mediante una vejiga natatoria
modificada.
Tenebrionidae.
Latreille,
1802.
|
 |
 |
|
Fragmentos de
sedimento con el molde negativo de un escarabajo
Tenebrionidae. |
Aspecto de un
Tenebrionidae en la actualidad, semejante al que vivió hace
miles de ańos. |
Artropodo Coleoptero.
El Museo
de Ciencias Naturales de Miramar dio a conocer el hallazgo de un
extrańo fósil atribuido a un escarabajo que habría vivido en la
región hace al menos 10.000 ańos antes del presente. Durante la
excavación, un fragmento de sedimento asociado a los huesos de un
perezoso gigante se partió en dos, revelando en ambas superficies
improntas con detalles muy finos de la morfología del escarabajo.
Solo se conservó el molde de la estructura original, ya que los
sedimentos fosilíferos de la región pampeana no preservan tejidos
blandos. Este tipo de hallazgos es extremadamente raro y aporta
valiosa información paleoambiental. La impronta permaneció
almacenada durante varios ańos debido a la ausencia de antecedentes
comparables que permitieran su identificación. Recién en 2012 se
realizaron estudios comparativos y los resultados fueron presentados
en 2014. El material fue identificado como perteneciente a la
familia Tenebrionidae, una de las más numerosas dentro de los
coleópteros, con unas 20.000 especies actuales, principalmente
asociadas a ambientes esteparios y desérticos. Estos escarabajos son
mayormente detritívoros, alimentándose de restos orgánicos en
descomposición de origen vegetal y animal, lo que sugiere que el
ejemplar hallado en Miramar se encontraba alimentándose de restos
orgánicos del cráneo del perezoso, como cuero y tendones. Por causas
desconocidas, el insecto murió junto a su fuente de alimento y se
preservó de manera excepcional durante miles de ańos, conservando
detalles morfológicos muy precisos. Los fósiles de insectos del
Cuaternario son escasos a nivel mundial; sin embargo, los pocos
conocidos suelen corresponder a especies actuales, sin evidencias
claras de extinciones o especiaciones durante este período, aunque
sí se registran importantes cambios en su distribución geográfica
asociados a variaciones climáticas.
Daimoniobarax
nephroides. Smith et al, 2011.
|
 |
|
 |
|
Aspecto de un
hormiguero fósil en Miramar. Foto del Museo de
Ciencias Naturales Punta Hermengo. |
|
Aspecto de una
comunidad de hormigas en forma ilustrativa (*). |
Insectos,
Formicidae.
En algunas zonas de la región pampeana se han
registrado estructuras biogénicas, es decir, formaciones generadas
por la actividad animal, identificadas como hormigueros fósiles.
Este icnofósil, asignado tentativamente al icnogénero
Daimoniobarax, está representado por sistemas subterráneos
de galerías con numerosas cámaras, algunas de ellas horizontales,
interconectadas mediante estrechos túneles y conductos verticales.
En las inmediaciones de Miramar, provincia de Buenos Aires, se han
documentado importantes concentraciones de icnofósiles de
hormigueros y termiteros correspondientes tanto al Plioceno como al
Pleistoceno. Las hormigas (Formicidae) constituyen una familia de
insectos eusociales que, al igual que avispas y abejas, pertenecen
al orden Hymenoptera. Su origen se remonta al Cretácico medio, hace
entre 110 y 130 millones de ańos, diversificándose notablemente tras
la expansión global de las plantas con flores.
Adelomelon beckii.
Broderip, 1836.
|
 |
|
 |
|
Adelomelon beckii,
del Pleistoceno de la localidad de Mar del Sud. Museo
de Ciencias Naturales de Miramar. |
|
Ejemplares de
Adelomelon brasiliana, de la formación Belgranense.
Museo de Cs Naturales de Miramar. |
Invertebrado.
Es un
gasterópodo de la familia Volutidae, caracterizado por presentar una
concha grande, ovalada y globosa, sólida y pesada, de color blanco
tiza en su superficie externa. Esta especie habita el fondo de la
plataforma marina, preferentemente en sustratos areno-fangosos. En
los yacimientos paleontológicos marinos, que afloran de manera poco
frecuente en el sudeste bonaerense, se han recuperado moluscos que
cuentan con representantes vivientes en el océano Atlántico y que
habitaron la región durante el Pleistoceno. Es muy inusual hallar
fósiles de origen marino en ambientes actualmente continentales,
fenómeno explicado por las variaciones del nivel del mar ocurridas a
lo largo de los milenios. Localmente, estos procesos se denominan
transgresiones o ingresiones marinas cuando el mar avanza sobre el
continente, y regresiones marinas cuando ocurre el fenómeno inverso.
Las primeras referencias paleontológicas sobre ingresiones marinas
en la región datan de mediados del siglo XIX y fueron realizadas por
d’Orbigny (1842), quien halló moluscos marinos en las cercanías de
la ciudad de Paraná y en la desembocadura del río Negro. Ańos más
tarde, Darwin (1846) registró la misma fauna en la península Valdés.
Otras especies asociadas incluyen Ostrea, Tagelus,
Mactra, foraminíferos (Ammonia, Elphidium),
ostrácodos (Cytheracea, Cyprideis), Adelomelon
brasiliana, Zidona sp., Pecten sp., Amiantis
purpurata, Glycymeris longior, Buccinanops sp.
y Olivancillaria sp.
Alternaria sp.
Nees, 1816.
|
 |
 |
 |
|
Espora de
hongo Alternaria sp, recuperada entre los huesos de
las patas de un Gliptodonte en la localidad de San Pedro. |
Presencia
característica de Alternaria sp, sobre una hoja en la
actualidad. |
Forma de
estudio (*). |
Paleobotanica, Hongos.
Es un hongo ascomiceto perteneciente al filo
Ascomycota. Las especies de este género constituyen algunos de los
principales patógenos de plantas y, en menor medida, pueden afectar
a animales mediante su ingesta. Alternaria es un
género fúngico muy común que incluye numerosas especies saprófitas,
endofíticas y patógenas, ampliamente distribuidas en el suelo y en
materia orgánica en descomposición. Estas especies sintetizan más de
70 metabolitos secundarios tóxicos para las plantas (fitotoxinas),
algunos de los cuales también afectan a animales, por lo que se
consideran micotoxinas. El Grupo Conservacionista de Fósiles del
Museo Paleontológico de San Pedro, junto al Centro de Ecología
Aplicada del Litoral (CONICET-UNNE), logró muestrear e identificar
granos de polen y esporas fósiles de distintas plantas y hongos que
habitaban los alrededores de un antiguo pantano cercano a la ciudad
de San Pedro, provincia de Buenos Aires, hace aproximadamente
700.000 ańos. El material fue recuperado entre las extremidades de
restos óseos de gliptodontes. El análisis de los fósiles y del
sedimento que los contenía permitió inferir que estos grandes
animales murieron en un antiguo humedal que actuaba como una “trampa
natural”, cuando se acercaban en busca de agua o alimento. Allí se
identificaron diminutos granos de polen y esporas fósiles
correspondientes a diversas familias de angiospermas, además de
esporas de hongos y restos de algas microscópicas, lo que indica un
ambiente de estepa herbácea sobre suelos arenosos, asociado a
cuerpos de agua cercanos.
|
(*) Imagen de autor u origen desconocido.
Ver sobre nuestras
Políticas de uso de imágenes.
|
|
|
Bibliografía Utilizada.
AGNOLIN Federico L., CHIMENTO Nicolás
R., CAMPO Denise H., MAGNUSSEN Mariano, BOH Daniel &
De CIANNI Francisco
(2018)
Large Carnivore Footprints from the Late Pleistocene of
Argentina, Ichnos,
ALBERDI, M.T.; CARTELLE, C. &
PRADO, J.L. 2003. El registro Pleistoceno de Equus
(Amerhippus) e Hippidion (Mammalia, Perissodactyla) de
Brasil. Consideraciones paleoecológicas y biosgeográficas.
Ameghiniana, 40:173-196.
ALBERDI, M.T.; CERDEŃO, E. &
PRADO, J.L. 2008. Stegomastodon platensis (Proboscidea,
Gomphotheriidae) en el Pleistoceno de Santiago del Estero,
Argentina. Ameghiniana, 45(2):257-271.
ALCARAZ, M.A.; FERRERO, B.S.
& NORIEGA, J.I. 2005. Primer registro de Antifer ultra
Ameghino, 1889 (Artiodactyla, Cervidae) en el Pleistoceno de
Entre Ríos. Revista del Instituto Superior de Correlación
Geología, INSUGEO, Miscelánea, 14:65-70.
AMEGHINO, F. 1888. Rápidas
diagnosis de mamíferos fósiles nuevos de la República
Argentina. Buenos Aires, Obras Completas, 5:471-480.
AMEGHINO, F. 1889.
Contribución al conocimiento de los mamíferos fósiles de la
República Argentina. Actas de la Academia Nacional de
Ciencias de Córdoba, 6:1-1027.
BERTON, Michael. (1992).
Dinosaurios y otros animales prehistóricos. Ediciones
Lrousse Argentina S.AI.C.
BERMAN, W.D. 1994. Los
carnívoros continentales (Mammalia, Carnivora) del Cenozoico
de la Provincia de Buenos Aires. Tesis Doctoral inédita,
Facultad de Ciencias Naturales y Museo, Universidad Nacional
de La Plata, La Plata, Argentina.
BONAPARTE, J.F. & BOBOVNIKOV, J. 1974.
Algunos fósiles pleistocénicos de la provincia de Tucumán y
su significado estratigráfico.
Acta Geológica Lilloana, 12(11):171-183.
BOGAN, S. & M.L. DE LOS
REYES. 2009. "Primer registro fósil del género Oligosarcus
GÜNTHER 1864 (Teleostei: Characiformes)"; Studia
Geologica Salmanticensia 45 (1): 41-52.
BOND, M.; CERDEŃO, E.P. & LOPEZ, G. 1995.
Los Ungulados Nativos de America del Sur. In:
Alberdi, M.T.; Leone, G. & Tonni, E.P. (Eds.), Evolución
climática y biológica de la región Pampeana durante los
últimos cinco millones de ańos. Un ensayo de correlación con
el Mediterráneo occidental. Museo Nacional de Ciencias
Naturales, Consejo Superior de Investigaciones Científicas,
Monografías 12, p. 259-275.
CABRERA, A. 1929. Sobre los
ciervos fósiles sudamericanos llamados Paraceros y
Morenelaphus. Memorias de la Real Sociedad Espańola de
Historia Natural, 15(1):53-64
CARLINI, A.A.; ZURITA, A.E. &
MIŃO-BOILINI, A.R. 2008. Reseńa paleobiogeográfica de los
Xenarthra (Mammalia) del Pleistoceno tardío de la región
Mesopotámica (Argentina). Revista del Instituto Superior de
Correlación Geológica, INSUGEO, 17:259-270.
CARTELLE GUERRA, C. &
BOHÓRQUEZ MAHECHA, G.A. 1985. Pampatherium paulacoutoi, uma
nova espécie de tatu gigante da Bahia, Brasil (Edentata,
Dasypodidae). Revista Brasileira de Zoologia, 2:229-254.
CIONE, A.L. & TONNI, E.P.
1995a. Bioestratigrafía y cronología del Cenozoico superior
de la región pampeana. In: Alberdi, M.T.; Leone, G. & Tonni,
E.P. (Eds.), Evolución climática y biológica de la región
Pampeana durante los últimos cinco millones de ańos. Un
ensayo de correlación con el Mediterráneo occidental. Museo
Nacional de Ciencias Naturales, Consejo Superior de
Investigaciones Científicas, Monografías, 12:47-74.
CRUZ, L. 2007. Xenarthra
(Mammalia) del Pleistoceno tardío-Holoceno temprano del
Departamento de Río Cuarto, provincia de Córdoba, Argentina.
Aspectos bioestratigráficos. Ameghiniana, 44(4):751-757.
ESTEBAN, G.I. 1996. Revisión
de los Mylodontinae cuaternarios (Edentata, Tardigrada) de
Argentina, Bolivia y Uruguay. Sistemática, Filogenia,
Paleobiología, Paleozoogeografía y Paleoecología. Tesis
Doctoral inédita, Instituto Miguel Lillo, Facultad de
Ciencias Naturales, Universidad Nacional de Tucumán, San
Miguel de Tucumán, Argentina.
FERRERO, B.S.; NORIEGA, J.I.
& ARAUJO, R. 2005. Primer hallazgo de Panthera onça Linnaeus
(Carnivora: Felidae) en el Pleistoceno tardío de la
provincia de Entre Ríos, Argentina. Ameghiniana, 42(4),
Resúmenes: 28R.
GASPARINI, G.M. & ZURITA,
A.E. 2005. Primer registro fósil de Tayassu pecari
(Mammalia, Artiodactyla) en la Argentina. Ameghiniana,
42:473-480.
GRAMAJO, A.J. 1992. La fauna
del Cuaternario y sus yacimientos en la Llanura Santiagueńa.
Museo de Ciencias Antropológicas y Naturales "Emilio y
Duncan Wagner", Serie Estudio, 4:75-93.
Holanda, Elizete Celestino, Brenda Soledad
Ferrero (2012) Reappraisal of the Genus Tapirus
(Perissodactyla, Tapiridae): Systematics and Phylogenetic
Affinities of the South American Tapirs.
Journal of
Mammalian Evolution marzo de 2012.
IRIONDO, M.H. 1994. Los
climas cuaternarios de la región pampeana. Comunicaciones
del Museo Provincial de Ciencias Naturales "Florentino
Ameghino", N.S., 4(2):1-46.
KRAGLIEVICH, J. 1932.
Contribución al conocimiento de los ciervos fósiles del
Uruguay. Anales del Museo de Historia Natural de Montevideo,
2(3):355-438.
LAZA, J. H. 1995. Signos de actividad de
insectos.
In: Alberdi, M.T.; Leone, G. & Tonni, E.P.
(eds.)
Evolución biológica y climática de la región
pampeana durante los últimos cinco millones de ańos, Un
ensayo de correlación con el Mediterráneo occidental. Museo
Nacional de Ciencias Naturales, Madrid, Monografías,
16:341–361.
MAGNUSSEN SAFFER, M. BOH, D Y ESTARLI,
C. 2014.
Asociación de un ejemplar juvenil y uno
adulto de Scelidotherium leptocephalum Owen, 1839
(Xenarthra, Mylodontidae, Scelidotheriinae ) en una
paleocueva en el Pleistoceno del Partido de General
Alvarado. Aspectos Sistemáticos y Tafonomicos.
XXVIII Jornadas Argentinas de Paleontología de Vertebrados.
Zapala – Villa El Chocon, Neuquén. Mayo del 2014.
Libro de Resúmenes.
, p 38.
MAGNUSSEN SAFFER, M. BOH, D Y ESTARLI,
C. 2014.
Primer registro de un
Tenebrionidae (Arthropoda, Coleoptera) asociado a restos
del cráneo de un Mylodontidae (Mammalia, Xenarthra),
en el Pleistoceno Superior de Punta Hermengo, Miramar,
Provincia de Buenos Aires, Republica Argentina. Libro de
resumenes.
III Jornadas
Paleontológicas del Centro.
MAGNUSSEN SAFFER, M.
2014.Perezosos gigantes
(Xenarthra,
Pilosa) del pleistoceno de la región pampeana. Paleo Revista
Argentina de PaleontoŞlogía. Boletín Paleontológico. Ańo
XII. 108: 08-13.
MAGNUSSEN SAFFER, M. BOH, D Y ESTARLI,
C. 2015.
Observaciones paleopatologicas y cambios morfológicos de
carácter funcional en una tibia de
Stegomastodon (Mammalia,
Gomphotheriidae) en el
Pleistoceno tardío del Partido de General Alvarado,
Provincia de Buenos Aires, Argentina.
XXIX Jornadas
Argentinas de Paleontología de Vertebrados.
Diamante, Entre Ríos. Mayo de 2015.
Libro de Resúmenes.
MAGNUSSEN SAFFER, M.
2016. Los meridiungulados característicos del Pleistoceno de
Argentina. Paleo Revista Argentina de Paleontología. Boletín
Paleontológico. Ańo XIV. 141: 17-21.
MAGNUSSEN MARIANO
&
BOH DANIEL. (2016).
Huellas de un tigre dientes de sable en el Pleistoceno
tardío de Miramar, provincia de Buenos Aires, República
Argentina. XXX Jornadas
Argentinas de Paleontología de Vertebrados. Buenos Aires.
Mayo de 2016. Libro de Resúmenes. Pag. 33.
MAGNUSSEN MARIANO
&
BOH DANIEL. (2018.
Las huellas fósiles de Miramar. Una historia de 100 mil
ańos. Museo Municipal Punta Hermengo. Serie divulgativa.
Segunda Edición.
MENEGAZ, A.N. & ORTIZ
JAUREGUIZAR, E. 1995. Los Artiodáctilos. In: Alberdi, M.T.;
Leone, G. & Tonni, E.P. (Eds.), Evolución climática y
biológica de la región Pampeana durante los últimos cinco
millones de ańos. Un ensayo de correlación con el
Mediterráneo occidental. Museo Nacional de Ciencias
Naturales, Consejo Superior de Investigaciones Científicas,
Monografías, 12:311-337.
NORIEGA, J.I.; CARLINI, A.A.
& TONNI, E.P. 2001. Vertebrados del Pleistoceno tardío de la
cuenca del arroyo Ensenada (Departamento Diamante, provincia
de Entre Ríos, Argentina). Bioestratigrafía y
paleobiogeografía. Ameghiniana, 38(4), Resúmenes: 38R.
NOVAS, F. 2006. Buenos Aires hace un millón
de ańos. Editorial Siglo XXI, Ciencia que Ladra. Serie
Mayor.
OLIVA CRISTIAN, DANIEL BOH, MARIANO MAGNUSSEN
&
CRISTIAN FAVIER DOBOIS.
(2018). Jornadas Regionales VI Arqueológicas y VII
Paleontológicas Miramar. Contribución al conocimiento de
Porcellusignum conculcator, Angulo y Casamiquela (Vertebratichnia,
Mammalipedia) del Cenozoico Superior (Mioceno Tardío –
Pleistoceno) de la Provincia de Buenos Aires (Argentina).
Libro de Resúmenes. Pag 33;34.
PARDIŃAS, U.F.J.
1995. Capítulo 11. Los roedores cricétidos. En: M.T.
Alberdi, G. Leone y E.P. Tonni (eds.) Evolución climática y
biológica de la región pampeana durante los últimos cinco
millones de ańos. Un ensayo de correlación con el
Mediterráneo occidental , Monografías del Museo Nacional de
Ciencias Naturales, Madrid, 14: 229-256.
PARDIŃAS, U.F.J.; GELFO, J.;
SAN CRISTÓBAL, J.; CIONE, A.L. & TONNI, E.P. 1996. Una
asociación de organismos marinos y continentales en el
Pleistoceno superior en el sur de la provincia de Buenos
Aires, Argentina. XIII Congreso Geológico Argentino and III
Congreso de Exploración de Hidrocarburos, Buenos Aires,
5:95-112.
PASCUAL, R.; ORTEGA HINOJOSA,
E.J.; GONDAR, D. & TONNI, E.P. 1966. Paleontografía
Bonaerense. Comisión de Investigaciones Científicas,
Provincia de Buenos Aires, 4:1-202.
PREVOSTI, F.J.
(2006). «Grandes cánidos (Carnivora, Canidae) del
Cuaternario de la República Argentina: sistemática,
filogenia, bioestratigrafía y paleoecología (tesis
doctoral)». Asociación Paleontológica Argentina: 501.
PREVOSTI,
F. J. 2006.
New materials of pleistocene cats (carnivora,
felidae) from southern South America, with comments on
biogeography and the fossil record.
Geobios, 39:
679-694.
REIG, O. (1980). Teoría del
origen y desarrollo de la fauna de mamíferos de América del
Sur. Mar del Plata, Museo Municipal de Ciencias Naturales
"Lorenzo Scaglia", 1981, Monografía Naturae, Nş 1.
RODRÍGUEZ-BUALÓ, S.; ZURITA,
A.E.; CARLINI, A.A. & FRIEDRICH, J. 2009. Los Pampatheriidae
(Xenarthra, Cingulata) de la provincia de Formosa,
Argentina. Un estudio sistemático y paleobiogeográfico
preliminar. Comunicaciones Científicas y Tecnológicas,
Universidad Nacional del Nordeste (UNNE).
SCILLATO-YANÉ, G.J. 1982. Los
Dasypodidae (Mammalia, Edentata) del Plioceno y Pleistoceno
de Argentina. Tesis Doctoral (inédita). Facultad de Ciencias
Naturales y Museo, Universidad Nacional de La Plata.
SOIBELZON,
LH. 2004. Revisión sistemática de los Tremarctinae
(Carnivora, Ursidae) fósiles de América del Sur. Revista del
Museo Argentino de Ciencias Naturales "Bernardino Rivadavia"
6(1): 107-133.
SOIBELZON,
LH; FI ISLA; A DONDAS. 2001. Primer registro de tres
individuos asociados de Arctotherium latidens (Ursidae:
Tremarctinae). Ameghiniana 38 (4)-Suplemento.
SOIBELZON,
E; GM
GASPARINI,
AE
ZURITA,
LH
SOIBELZON.
2008. Las “toscas del Río de La
Plata” (Buenos Aires, Argentina). Una actualización
paleofaunística. Revista del Museo Argentino de Ciencias
Naturales "Bernardino Rivadavia" 19(2): 291-308.
SOIBELZON, L.H.; ZURITA A.E.;
MORGAN, C.C.; RODRÍGUEZ, S.; GASPARINI, G.M.; SOIBELZON, E.;
SCHUBERT, B.W. & MIŃO-BOILINI, A.R. 2010. Primer registro
fósil de Procyon cancrivorus (G. Cuvier, 1798) (Carnivora,
Procyonidae) en la Argentina. Revista Mexicana de Ciencias
Geológicas, 27(2):313-319
TAMBUSSI, C. P., 1995. Aves. En: ALBERDI, M.
T., G. LEONE y E. P. TONNI (eds.), Evolución biológica y
climática de la Región Pampeana durante los últimos 5
millones de ańos. Monografía del Museo Nacional de Ciencias
Naturales, CSIC, Espańa, cap. 7: 145-161.
TASSARA, D.A.; SCIAN, R.D. & OSTERRIETH, M.
2005.
Hallazgo de paleocuevas en área costera de
las localidades de Camet Norte y Mar de Cobo (partido de Mar
Chiquita), provincia de Buenos Aires. IV Jornadas
Paleontológicas Regionales, Actas, p. 16.
TONNI, E.P. & FIDALGO, F.
1978. Consideraciones sobre los cambios climáticos durante
el Pleistoceno tardío-Reciente en la provincia de Buenos
Aires. Aspectos ecológicos y zoogeográficos relacionados.
Ameghiniana, 15(1-2):235-253
TONNI, E. P. Y FIDALGO, F. 1982. Geología y
Paleontología de los sedimentos del Pleistoceno en el área
de Punta Hermengo (Miramar, prov.
Bs. As, Repub.
Argentina); Aspectos paleoclimaticos.
Ameghiniana 19 (1-2): 79-108.
TONNI, E.P. 2004. Faunas y
Climas en el cuaternario de la Mesopotamia Argentina. In:
Aceńolaza, R.G. (Coord.). Temas de la Biodiversidad del
Litoral fluvial argentino, INSUGEO, Miscelánea, 12:31-36.
Zacarías, G.G., De La Fuente,
M. S., Fernández, M. S. y Zurita, A. E.2013 Nueva especie se
tortuga terrestre gigante del Género Chelonoidis fitzinger,
1835 (Cryptodira: Testudinidae), del Miembro Inferior de la
Formación Toropí/ Yupoí (Pleistoceno Tardío/ Lujanense),
Bella Vista, Corrientes, Argentina. Ameghiniana 50 (3): 298
– 318.
ZARATE, M.A. 1989.
Estratigrafía y geología del CenozoicoTardío en los
acantilados marinos comprendidosentre Playa San Carlos y
Arroyo Chapadmalal. Pdo. Gral. Pueyrredón, Prov. de Bs. As.
Facultad de Ciencias Naturales y Museo, Universidad Nacional
de La Plata, Tesis Doctoral, 221 p.
Ver mas bibliografía utilizada para hacer la presente
Pagina;
AQUÍ.
Ver bibliografía de divulgación sugerida;
AQUÍ. |
|
|
|
fosiles
Pleistoceno fosiles Pleistoceno fosiles Pleistoceno fosiles
Pleistoceno fosiles Pleistoceno fosiles Pleistoceno fosiles |
    |
 Ver otros
continentes en el mismo Periodo Geológico
Ver otros
continentes en el mismo Periodo Geológico |
|